

Peak and persistent excess of genetic diversity following an abrupt migration increase
- PMID: 23307901
- PMCID: PMC3584009
- DOI: 10.1534/genetics.112.147785
Peak and persistent excess of genetic diversity following an abrupt migration increase
Abstract
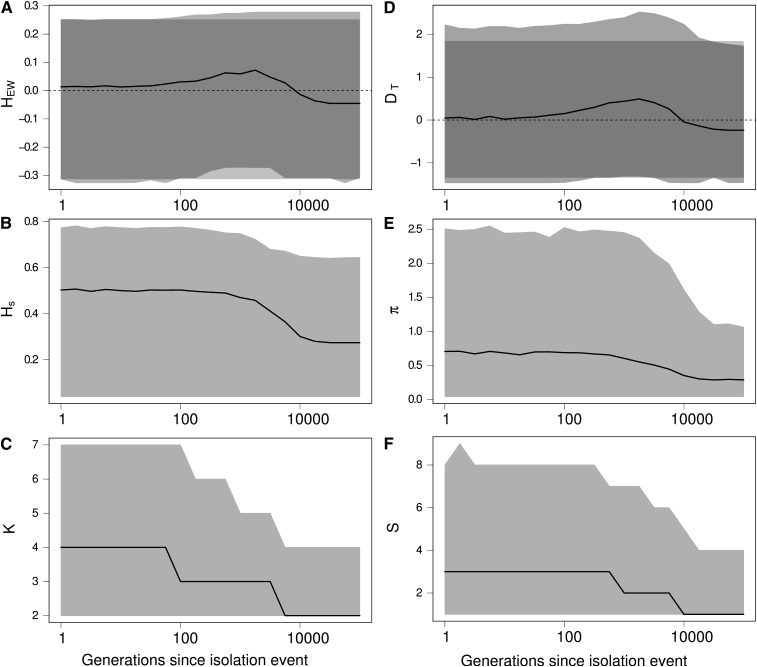
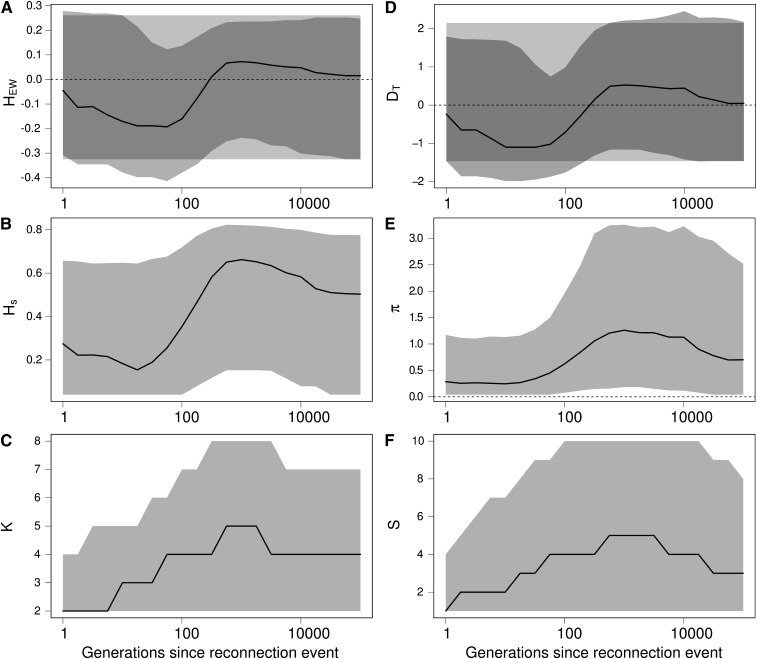
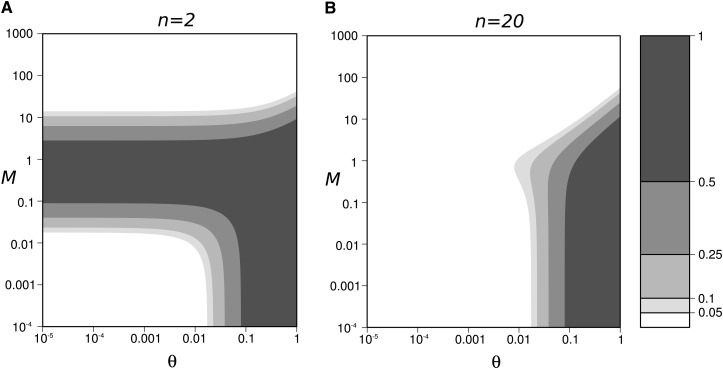
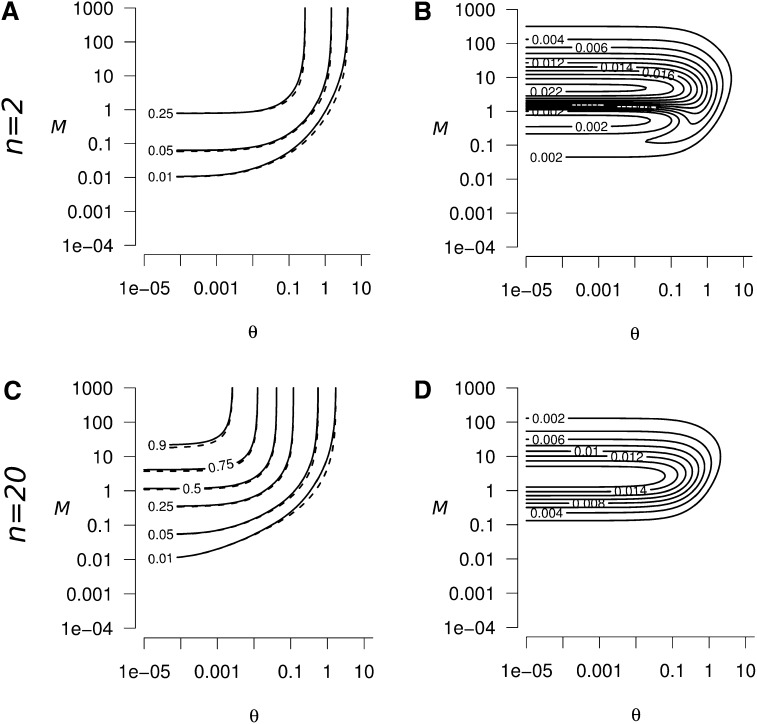
Genetic diversity is essential for population survival and adaptation to changing environments. Demographic processes (e.g., bottleneck and expansion) and spatial structure (e.g., migration, number, and size of populations) are known to shape the patterns of the genetic diversity of populations. However, the impact of temporal changes in migration on genetic diversity has seldom been considered, although such events might be the norm. Indeed, during the millions of years of a species' lifetime, repeated isolation and reconnection of populations occur. Geological and climatic events alternately isolate and reconnect habitats. We analytically document the dynamics of genetic diversity after an abrupt change in migration given the mutation rate and the number and sizes of the populations. We demonstrate that during transient dynamics, genetic diversity can reach unexpectedly high values that can be maintained over thousands of generations. We discuss the consequences of such processes for the evolution of species based on standing genetic variation and how they can affect the reconstruction of a population's demographic and evolutionary history from genetic data. Our results also provide guidelines for the use of genetic data for the conservation of natural populations.
Figures
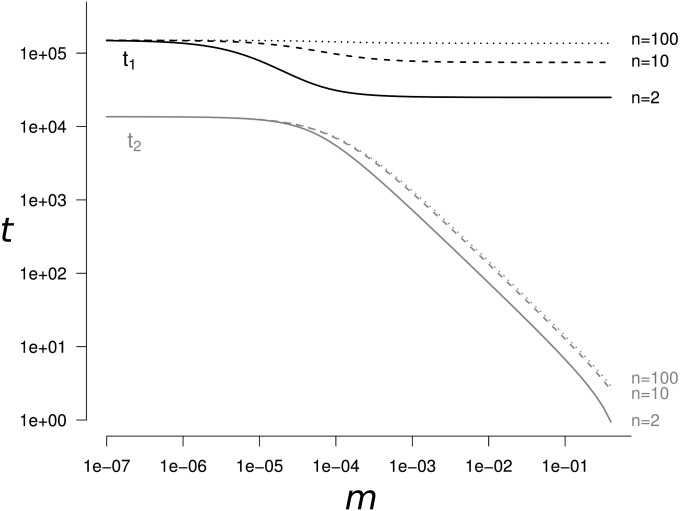
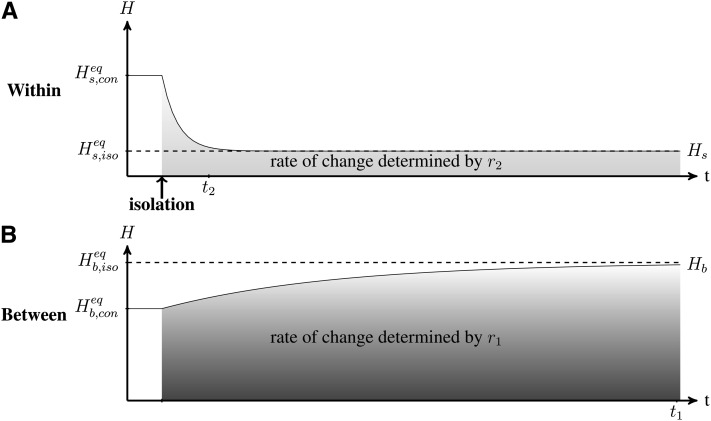
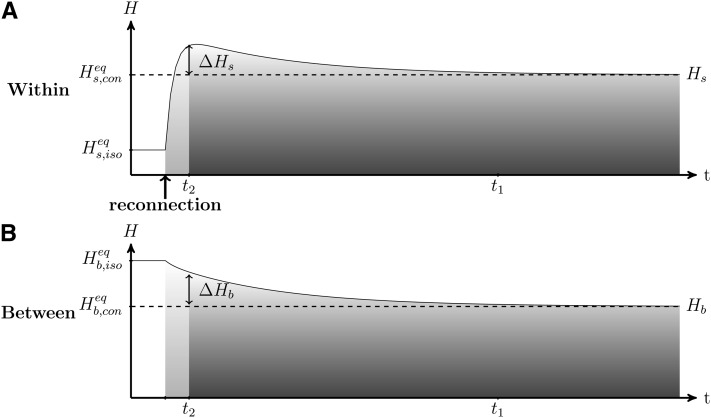
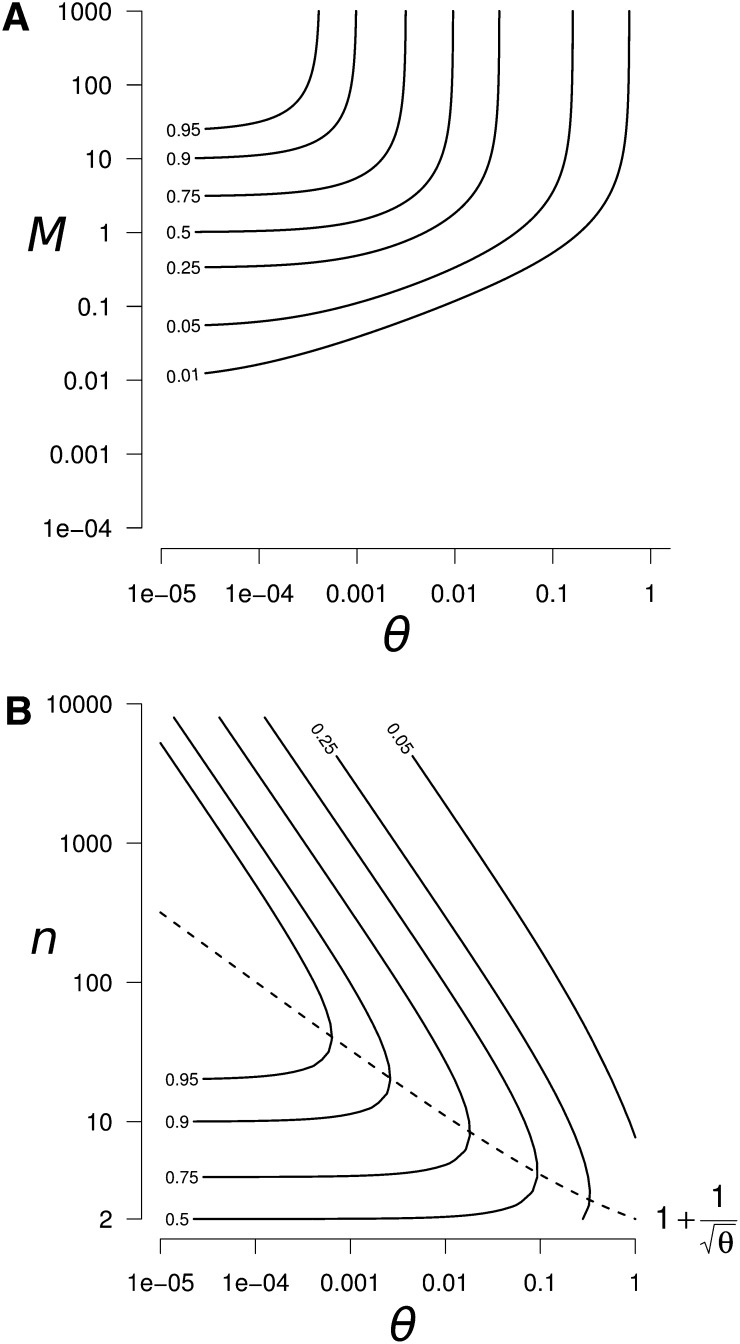
References
-
- Antonelli A., Sanmartín I., 2011. Why are there so many plant species in the neotropics? Taxon 60: 403–414.
-
- Arnegard M. E., Markert J. A., Danley P. D., Stauffer J. R., Ambali A. J., et al. , 1999. Population structure and colour variation of the cichlid fishes labeotropheus fuelleborni ahl along a recently formed archipelago of rocky habitat patches in southern Lake Malawi. Proc. R. Soc. Lond. B Biol. Sci. 266: 119–130.
-
- Barrett R. D. H., Schluter D., 2008. Adaptation from standing genetic variation. Trends Ecol. Evol. 23: 38–44. - PubMed
-
- Barrier M., Baldwin B. G., Robichaux R. H., Purugganan M. D., 1999. Interspecific hybrid ancestry of a plant adaptive radiation: allopolyploidy of the Hawaiian silversword alliance (Asteraceae) inferred from floral homeotic gene duplications. Mol. Biol. Evol. 16: 1105–1113. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
