

Evidence for horizontal transmission of secondary endosymbionts in the Bemisia tabaci cryptic species complex
- PMID: 23308142
- PMCID: PMC3538644
- DOI: 10.1371/journal.pone.0053084
Evidence for horizontal transmission of secondary endosymbionts in the Bemisia tabaci cryptic species complex
Abstract
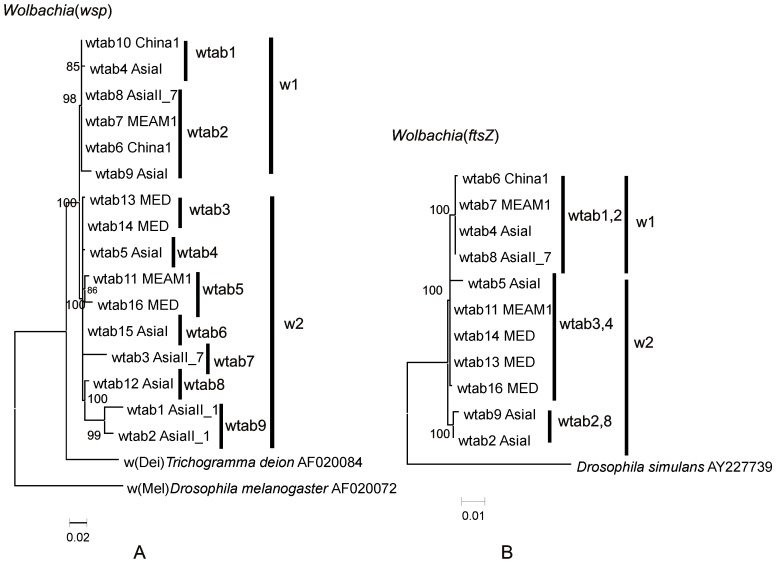
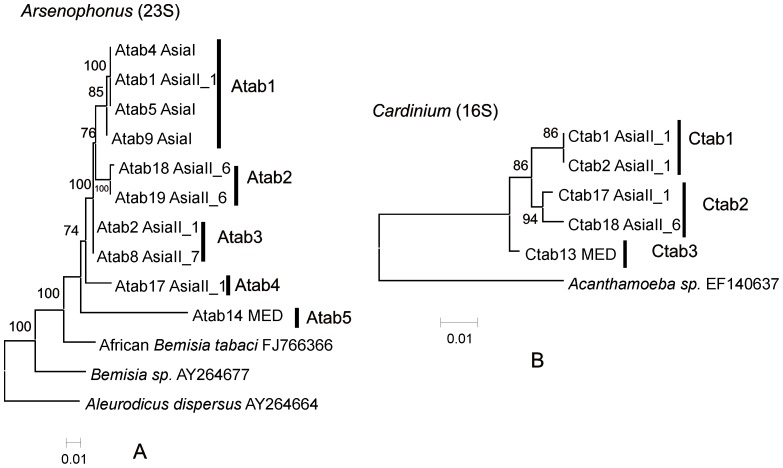
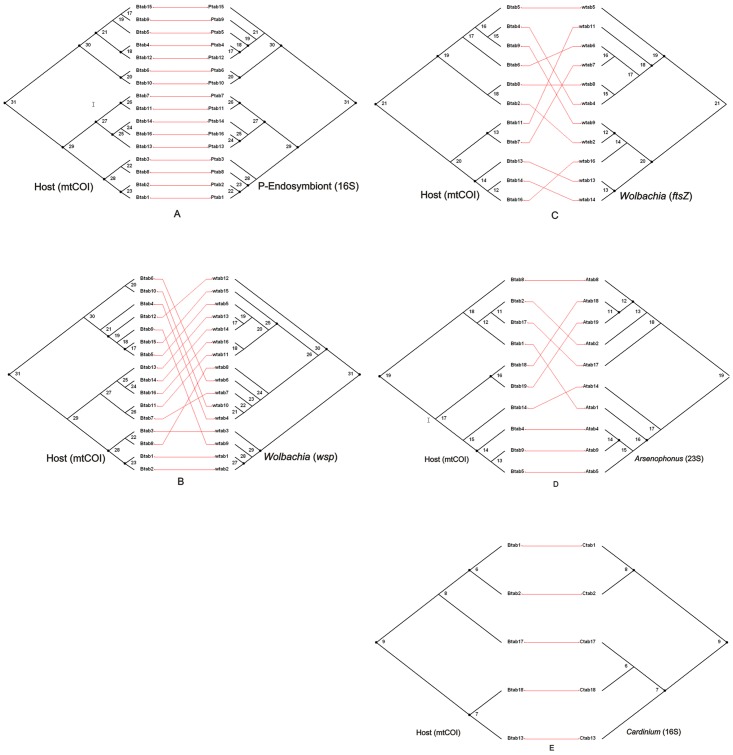
Bemisia tabaci (Hemiptera: Aleyrodidae) is a globally distributed pest composed of at least 34 morphologically indistinguishable cryptic species. At least seven species of endosymbiont have been found infecting some or all members of the complex. The origin(s) of the associations between specific endosymbionts and their whitefly hosts is unknown. Infection is normally vertical, but horizontal transmission does occur and is one way for new infections to be introduced into individuals. The relationships between the different members of the cryptic species complex and the endosymbionts have not been well explored. In this study, the phylogenies of different cryptic species of the host with those of their endosymbionts were compared. Of particular interest was whether there was evidence for both coevolution and horizontal transmission. Congruence was observed for the primary endosymbiont, Portiera aleyrodidarum, and partial incongruence in the case of two secondary endosymbionts, Arsenophonus and Cardinium and incongruence for a third, Wolbachia. The patterns observed for the primary endosymbiont supported cospeciation with the host while the patterns for the secondary endosymbionts, and especially Wolbachia showed evidence of host shifts and extinctions through horizontal transmission rather than cospeciation. Of particular note is the observation of several very recent host shift events in China between exotic invader and indigenous members of the complex. These shifts were from indigenous members of the complex to the invader as well as from the invader to indigenous relatives.
Conflict of interest statement
Figures

References
-
- Moran NA, Baumann P (2000) Bacterial endosymbionts in animals. Curr Opin Microbiol 3: 270–275. - PubMed
-
- Baumann P (2005) Biology of bacteriocyte-associated endosymbionts of plant sap-sucking insects. Ann Rev Microbiol 59: 155–189. - PubMed
-
- Moran NA, Telang A (1998) Bacteriocyte-associated symbionts of insects: a variety of insect groups harbor ancient prokaryotic endosymbionts. BioScience 48: 295–304.
-
- Buchner P (1965) Endosymbiosis of animals with plant microorganisms, p. 332–338. John Wiley and Sons Interscience, New York, N.Y.
-
- Costa HS, Westcot DM, Ullman DE, Rosell R, Brown JK, et al. (1995) Morphological variation in Bemisia endosymbionts. Protoplasma 189: 194–202.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
