

Fast silencing reveals a lost role for reciprocal inhibition in locomotion
- PMID: 23312521
- PMCID: PMC3542422
- DOI: 10.1016/j.neuron.2012.10.040
Fast silencing reveals a lost role for reciprocal inhibition in locomotion
Abstract
Alternating contractions of antagonistic muscle groups during locomotion are generated by spinal "half-center" networks coupled in antiphase by reciprocal inhibition. It is widely thought that reciprocal inhibition only coordinates the activity of these muscles. We have devised two methods to rapidly and selectively silence neurons on just one side of Xenopus tadpole spinal cord and hindbrain, which generate swimming rhythms. Silencing activity on one side led to rapid cessation of activity on the other side. Analyses reveal that this resulted from the depression of reciprocal inhibition connecting the two sides. Although critical neurons in intact tadpoles are capable of pacemaker firing individually, an effect that could support motor rhythms without inhibition, the swimming network itself requires ~23 min to regain rhythmic activity after blocking inhibition pharmacologically, implying some homeostatic changes. We conclude therefore that reciprocal inhibition is critical for the generation of normal locomotor rhythm.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
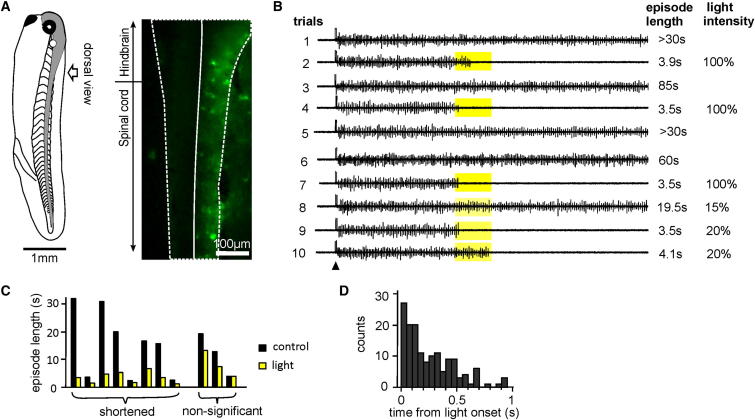
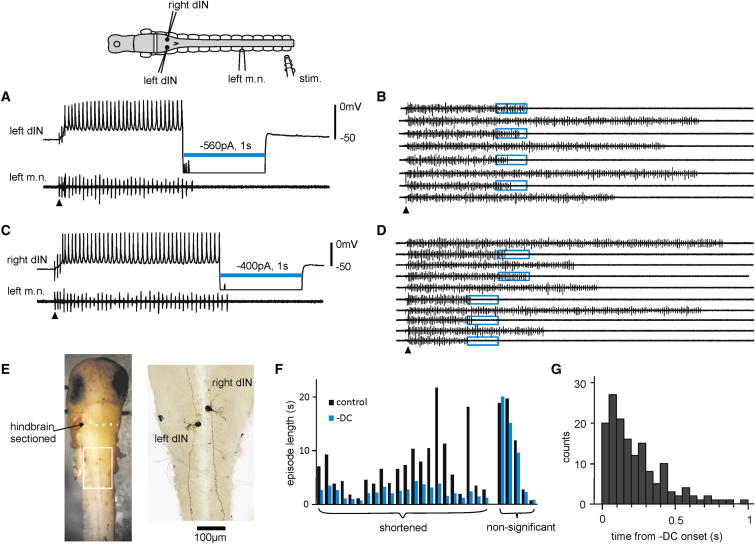
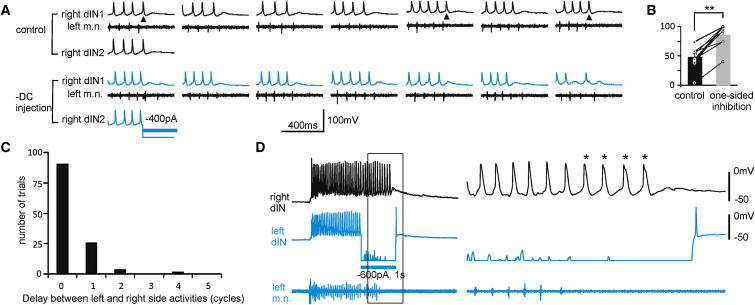
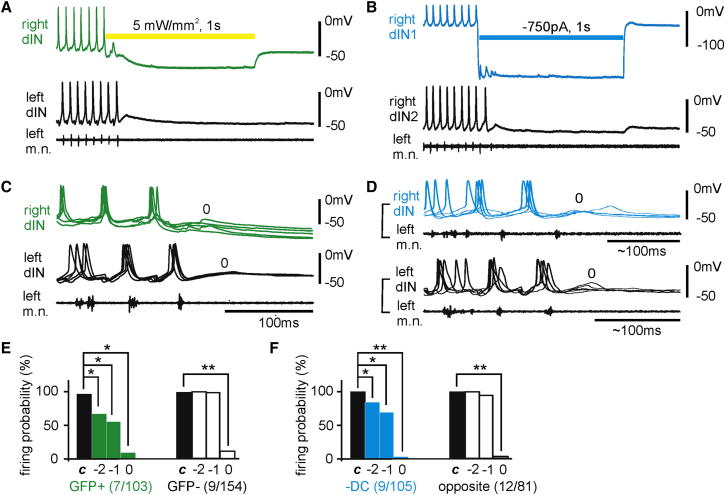
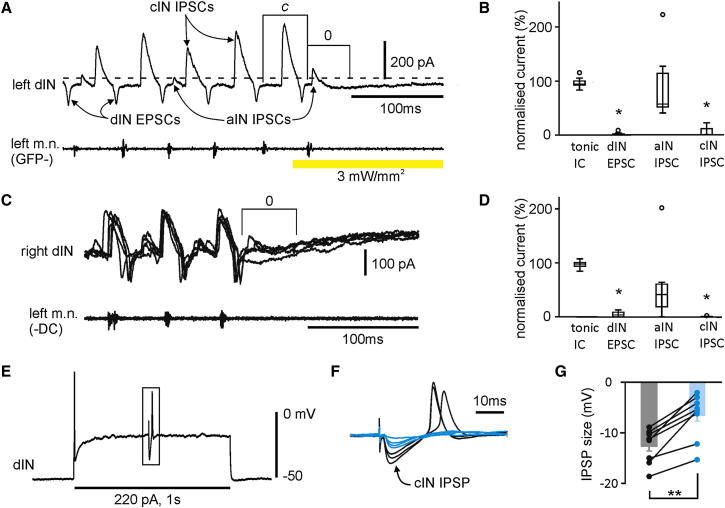
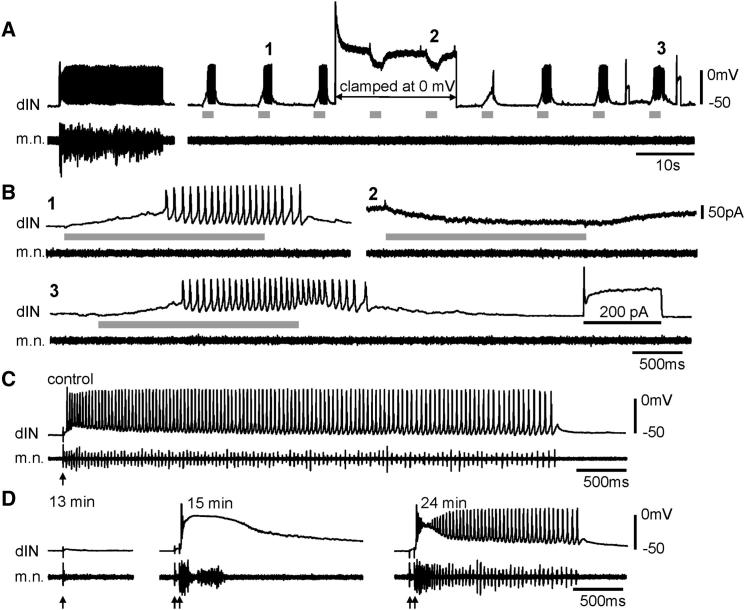
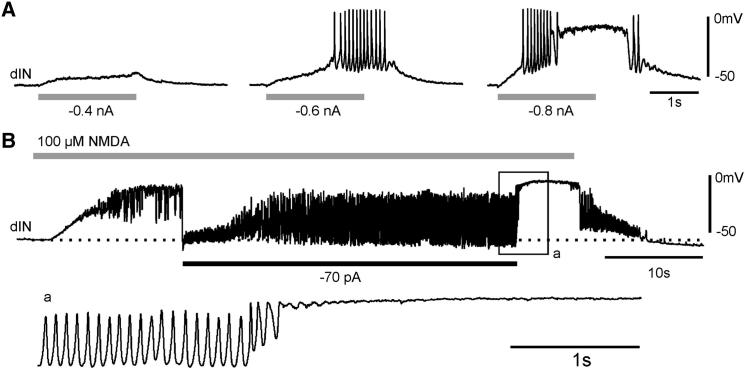
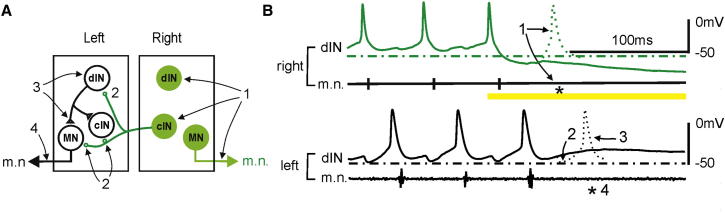
Similar articles
-
Generation of locomotion rhythms without inhibition in vertebrates: the search for pacemaker neurons.Integr Comp Biol. 2011 Dec;51(6):879-89. doi: 10.1093/icb/icr021. Epub 2011 May 11. Integr Comp Biol. 2011. PMID: 21562024
-
Specific brainstem neurons switch each other into pacemaker mode to drive movement by activating NMDA receptors.J Neurosci. 2010 Dec 8;30(49):16609-20. doi: 10.1523/JNEUROSCI.3695-10.2010. J Neurosci. 2010. PMID: 21148000 Free PMC article.
-
The generation of antiphase oscillations and synchrony by a rebound-based vertebrate central pattern generator.J Neurosci. 2014 Apr 23;34(17):6065-77. doi: 10.1523/JNEUROSCI.4198-13.2014. J Neurosci. 2014. PMID: 24760866 Free PMC article.
-
Positive feedback as a general mechanism for sustaining rhythmic and non-rhythmic activity.J Physiol Paris. 1995;89(4-6):241-8. doi: 10.1016/0928-4257(96)83640-0. J Physiol Paris. 1995. PMID: 8861822 Review.
-
Roles for inhibition: studies on networks controlling swimming in young frog tadpoles.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2008 Feb;194(2):185-93. doi: 10.1007/s00359-007-0273-3. Epub 2008 Jan 29. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2008. PMID: 18228081 Review.
Cited by
-
Modelling the Effects of Electrical Coupling between Unmyelinated Axons of Brainstem Neurons Controlling Rhythmic Activity.PLoS Comput Biol. 2015 May 8;11(5):e1004240. doi: 10.1371/journal.pcbi.1004240. eCollection 2015 May. PLoS Comput Biol. 2015. PMID: 25954930 Free PMC article.
-
Mechanosensory Stimulation Evokes Acute Concussion-Like Behavior by Activating GIRKs Coupled to Muscarinic Receptors in a Simple Vertebrate.eNeuro. 2017 Apr 27;4(2):ENEURO.0073-17.2017. doi: 10.1523/ENEURO.0073-17.2017. eCollection 2017 Mar-Apr. eNeuro. 2017. PMID: 28462392 Free PMC article.
-
The CPGs for Limbed Locomotion-Facts and Fiction.Int J Mol Sci. 2021 May 30;22(11):5882. doi: 10.3390/ijms22115882. Int J Mol Sci. 2021. PMID: 34070932 Free PMC article. Review.
-
Bursting emerges from the complementary roles of neurons in a four-cell network.J Neurophysiol. 2022 Apr 1;127(4):1054-1066. doi: 10.1152/jn.00017.2022. Epub 2022 Mar 23. J Neurophysiol. 2022. PMID: 35320029 Free PMC article.
-
Selective Gating of Neuronal Activity by Intrinsic Properties in Distinct Motor Rhythms.J Neurosci. 2015 Jul 8;35(27):9799-810. doi: 10.1523/JNEUROSCI.0323-15.2015. J Neurosci. 2015. PMID: 26156983 Free PMC article.
References
-
- Arshavsky Yu.I., Orlovsky G.N., Panchin Yu.V., Roberts A., Soffe S.R. Neuronal control of swimming locomotion: analysis of the pteropod mollusc Clione and embryos of the amphibian Xenopus. Trends Neurosci. 1993;16:227–233. - PubMed
-
- Beato M., Nistri A. Interaction between disinhibited bursting and fictive locomotor patterns in the rat isolated spinal cord. J. Neurophysiol. 1999;82:2029–2038. - PubMed
-
- Brown T.G. The intrinsic factors in the act of progression in the mammal. Proc. R. Soc. Lond. B. 1911;84:308–319.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
