

The grape berry-specific basic helix-loop-helix transcription factor VvCEB1 affects cell size
- PMID: 23314819
- PMCID: PMC3580811
- DOI: 10.1093/jxb/ers374
The grape berry-specific basic helix-loop-helix transcription factor VvCEB1 affects cell size
Abstract
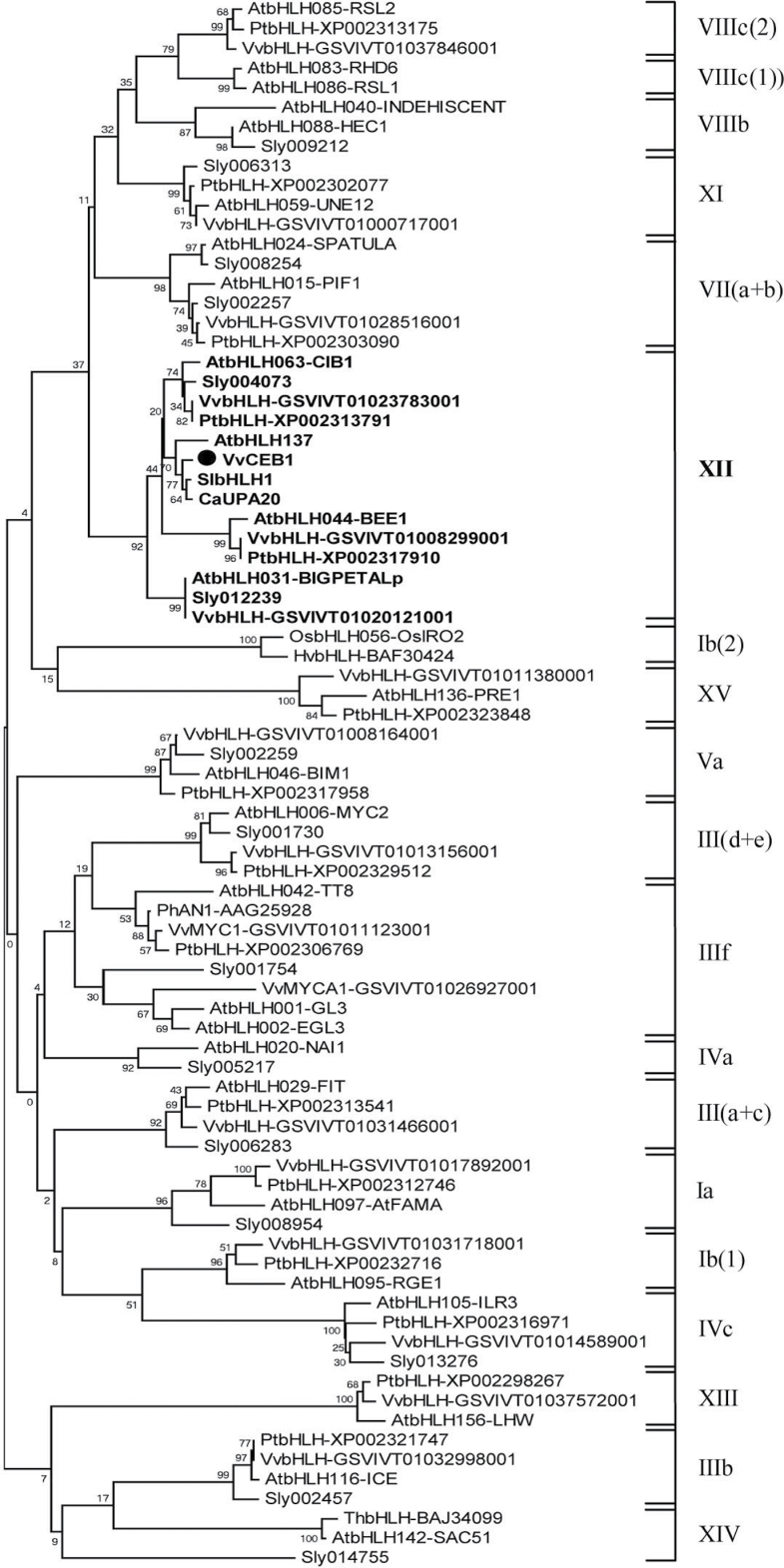
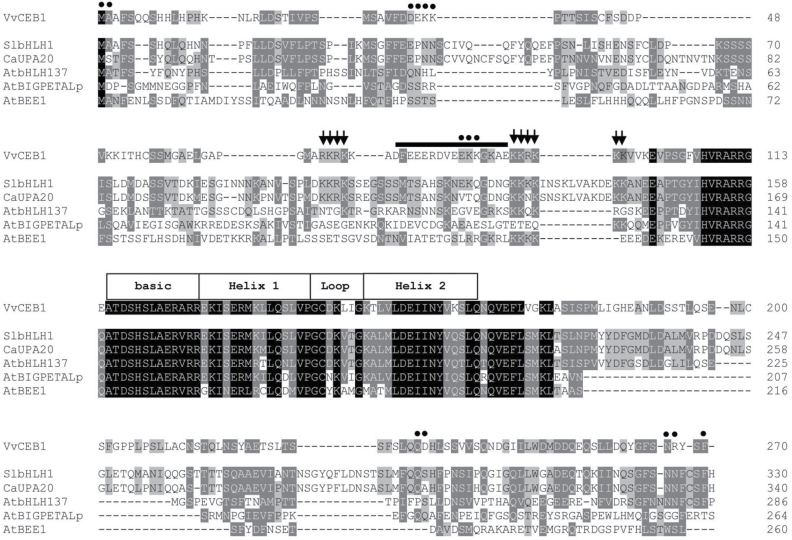
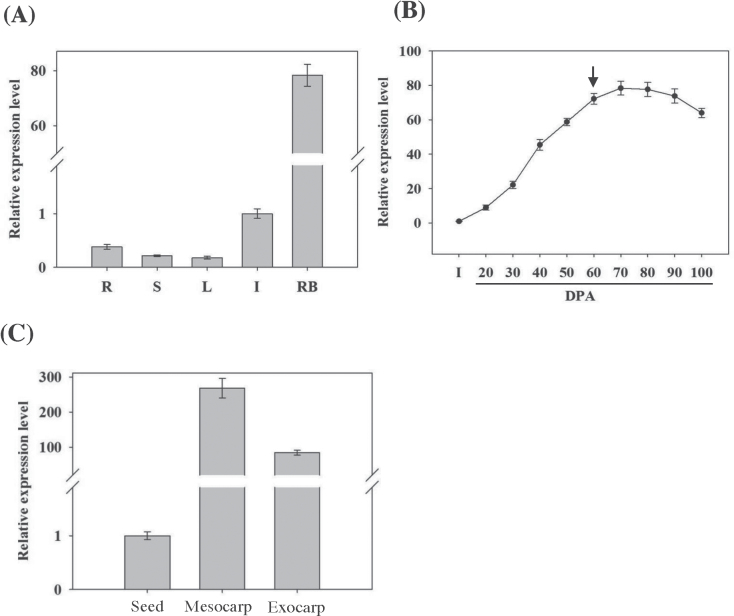
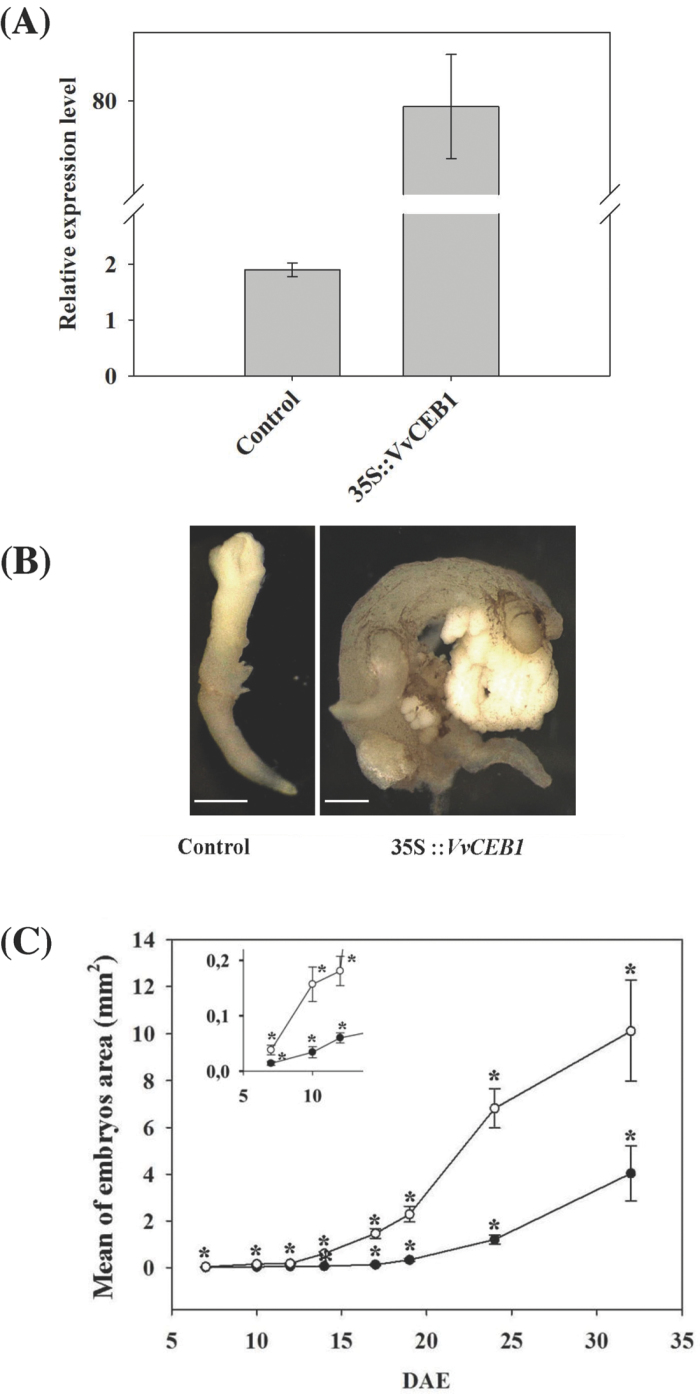
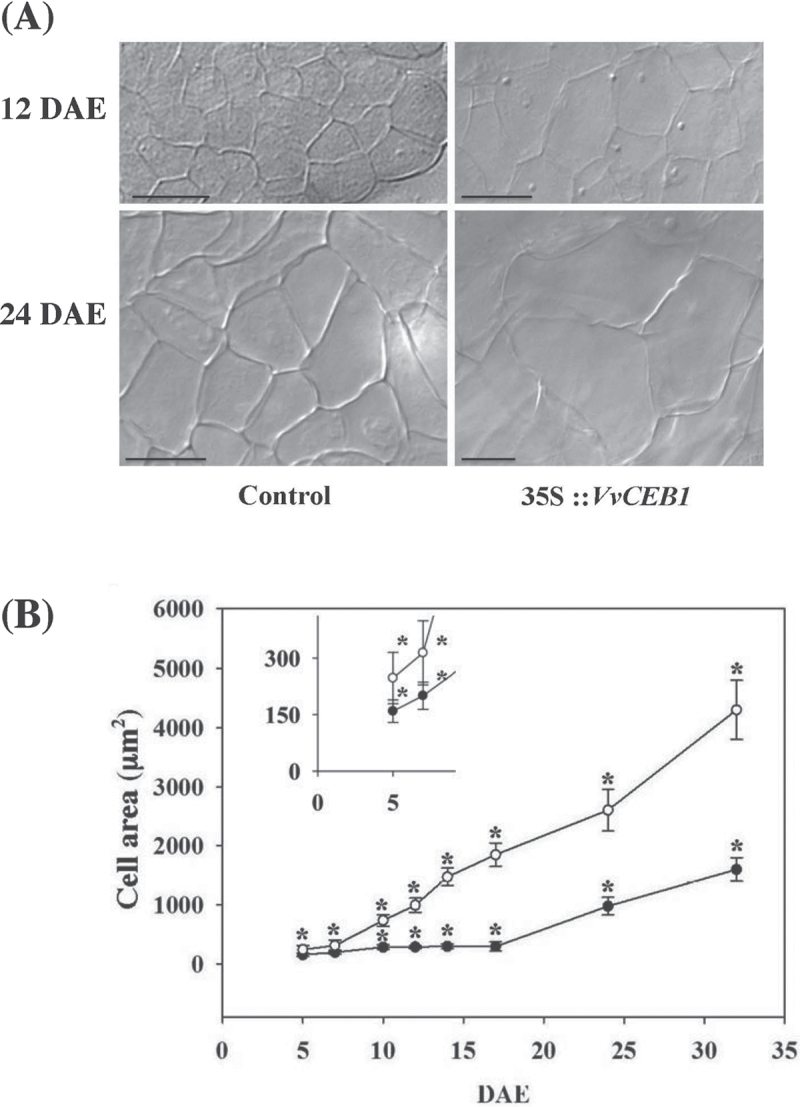
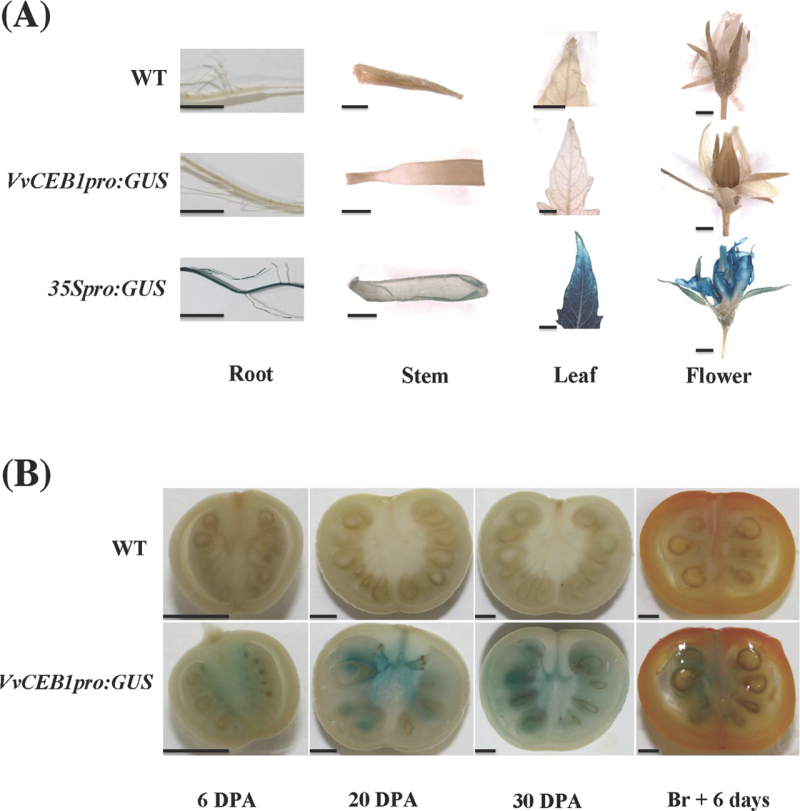
The development of fleshy fruits involves complex physiological and biochemical changes. After fertilization, fruit growth usually begins with cell division, continues with both cell division and expansion, allowing fruit set to occur, and ends with cell expansion only. In spite of the economical importance of grapevine, the molecular mechanisms controlling berry growth are not fully understood. The present work identified and characterized Vitis vinifera cell elongation bHLH protein (VvCEB1), a basic helix-loop-helix (bHLH) transcription factor controlling cell expansion in grape. VvCEB1 was expressed specifically in berry-expanding tissues with a maximum around veraison. The study of VvCEB1 promoter activity in tomato confirmed its specific fruit expression during the expansion phase. Overexpression of VvCEB1 in grape embryos showed that this protein stimulates cell expansion and affects the expression of genes involved in cell expansion, including genes of auxin metabolism and signalling. Taken together, these data show that VvCEB1 is a fruit-specific bHLH transcription factor involved in grape berry development.
Figures

References
-
- Berleth T, Krogan NT, Scarpella E. 2004. Auxin signals—turning genes on and turning cells around. Current Opinion in Plant Biology 7, 553–563 - PubMed
-
- Blouin J, Guimberteau G. 2000. Maturation et maturité des raisins. France:Féret;
-
- Böttcher C, Boss PK, Davies C. 2011. Acyl substrate preferences of an IAA-amido synthetase account for variations in grape (Vitis vinifera L.) berry ripening caused by different auxinic compounds indicating the importance of auxin conjugation in plant development. Journal of Experimental Botany 62, 4267–4280 - PMC - PubMed
-
- Carrari F, Fernie AR. 2006. Metabolic regulation underlying tomato fruit development. Journal of Experimental Botany 57, 1883–1897 - PubMed
-
- Cartharius K, Frech K, Grote K, Klocke B, Haltmeier M, Klingenhoff A, Frisch M, Bayerlein M, Werner T. 2005. MatInspector and beyond: promoter analysis based on transcription factor binding sites. Bioinformatics 21, 2933–2942 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
