

Characterization of four new mouse cytochrome P450 enzymes of the CYP2J subfamily
- PMID: 23315644
- PMCID: PMC3608456
- DOI: 10.1124/dmd.112.049429
Characterization of four new mouse cytochrome P450 enzymes of the CYP2J subfamily
Abstract
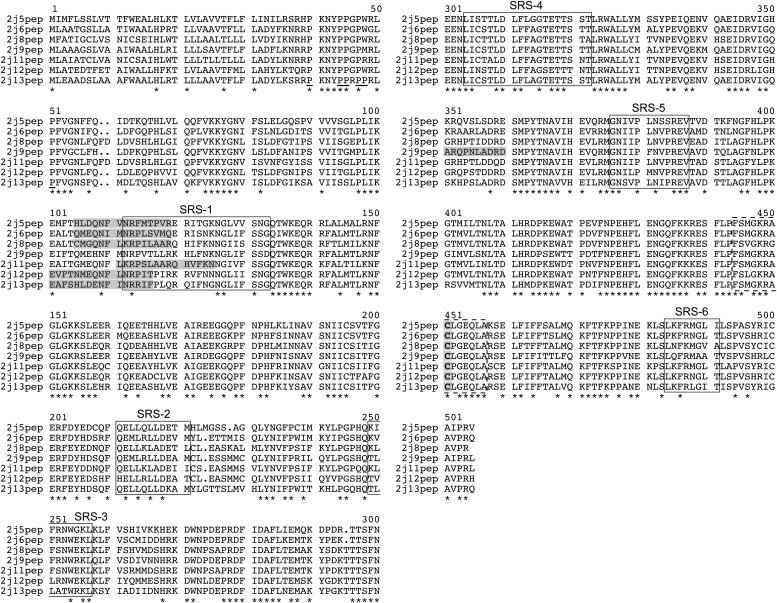
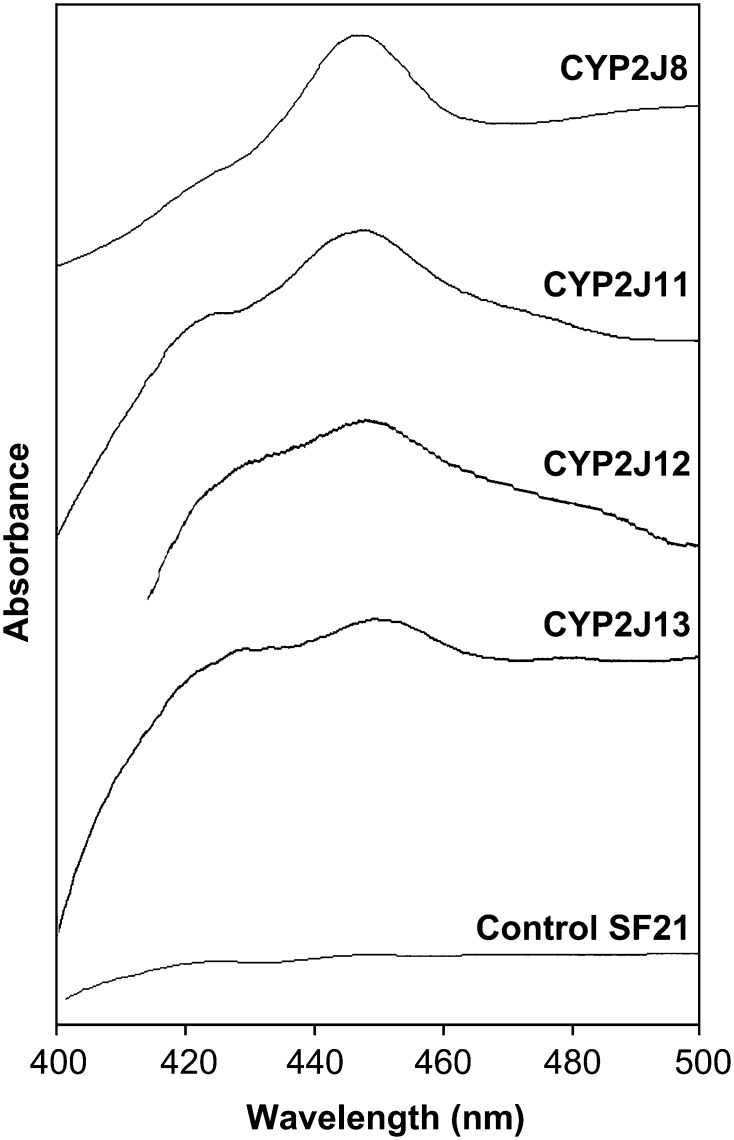
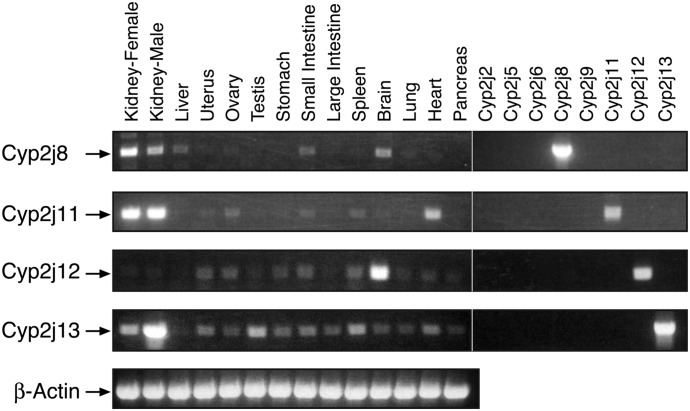
The cytochrome P450 superfamily encompasses a diverse group of enzymes that catalyze the oxidation of various substrates. The mouse CYP2J subfamily includes members that have wide tissue distribution and are active in the metabolism of arachidonic acid (AA), linoleic acid (LA), and other lipids and xenobiotics. The mouse Cyp2j locus contains seven genes and three pseudogenes located in a contiguous 0.62 megabase cluster on chromosome 4. We describe four new mouse CYP2J isoforms (designated CYP2J8, CYP2J11, CYP2J12, and CYP2J13). The four cDNAs contain open reading frames that encode polypeptides with 62-84% identity with the three previously identified mouse CYP2Js. All four new CYP2J proteins were expressed in Sf21 insect cells. Each recombinant protein metabolized AA and LA to epoxides and hydroxy derivatives. Specific antibodies, mRNA probes, and polymerase chain reaction primer sets were developed for each mouse CYP2J to examine their tissue distribution. CYP2J8 transcripts were found in the kidney, liver, and brain, and protein expression was confirmed in the kidney and brain (neuropil). CYP2J11 transcripts were most abundant in the kidney and heart, with protein detected primarily in the kidney (proximal convoluted tubules), liver, and heart (cardiomyocytes). CYP2J12 transcripts were prominently present in the brain, and CYP2J13 transcripts were detected in multiple tissues, with the highest expression in the kidney. CYP2J12 and CYP2J13 protein expression could not be determined because the antibodies developed were not immunospecific. We conclude that the four new CYP2J isoforms might be involved in the metabolism of AA and LA to bioactive lipids in mouse hepatic and extrahepatic tissues.
Figures




References
-
- Bylund J, Kunz T, Valmsen K, Oliw EH. (1998) Cytochromes P450 with bisallylic hydroxylation activity on arachidonic and linoleic acids studied with human recombinant enzymes and with human and rat liver microsomes. J Pharmacol Exp Ther 284:51–60 - PubMed
-
- Campbell WB, Gebremedhin D, Pratt PF, Harder DR. (1996) Identification of epoxyeicosatrienoic acids as endothelium-derived hyperpolarizing factors. Circ Res 78:415–423 - PubMed
-
- Campbell WB, Harder DR. (1999) Endothelium-derived hyperpolarizing factors and vascular cytochrome P450 metabolites of arachidonic acid in the regulation of tone. Circ Res 84:484–488 - PubMed
-
- Capdevila JH. (2007) Regulation of ion transport and blood pressure by cytochrome p450 monooxygenases. Curr Opin Nephrol Hypertens 16:465–470 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
