

Reelin induces EphB activation
- PMID: 23318582
- PMCID: PMC3616423
- DOI: 10.1038/cr.2013.7
Reelin induces EphB activation
Abstract
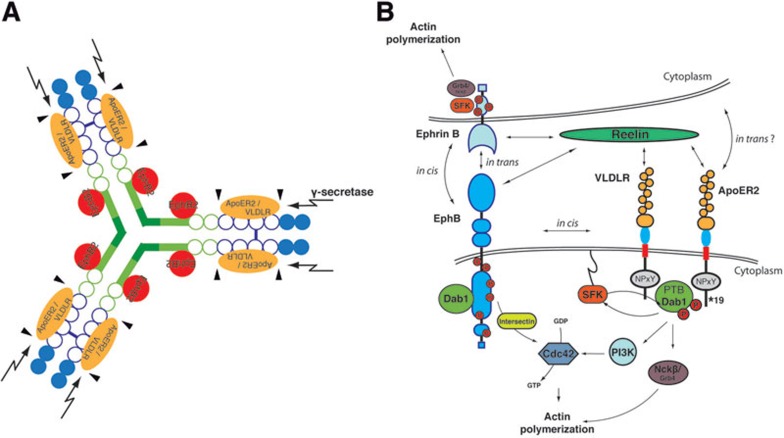
The integration of newborn neurons into functional neuronal networks requires migration of cells to their final position in the developing brain, the growth and arborization of neuronal processes and the formation of synaptic contacts with other neurons. A central player among the signals that coordinate this complex sequence of differentiation events is the secreted glycoprotein Reelin, which also modulates synaptic plasticity, learning and memory formation in the adult brain. Binding of Reelin to ApoER2 and VLDL receptor, two members of the LDL receptor family, initiates a signaling cascade involving tyrosine phosphorylation of the intracellular cytoplasmic adaptor protein Disabled-1, which targets the neuronal cytoskeleton and ultimately controls the positioning of neurons throughout the developing brain. However, it is possible that Reelin signals interact with other receptor-mediated signaling cascades to regulate different aspects of brain development and plasticity. EphB tyrosine kinases regulate cell adhesion and repulsion-dependent processes via bidirectional signaling through ephrin B transmembrane proteins. Here, we demonstrate that Reelin binds to the extracellular domains of EphB transmembrane proteins, inducing receptor clustering and activation of EphB forward signaling in neurons, independently of the 'classical' Reelin receptors, ApoER2 and VLDLR. Accordingly, mice lacking EphB1 and EphB2 display a positioning defect of CA3 hippocampal pyramidal neurons, similar to that in Reelin-deficient mice, and this cell migration defect depends on the kinase activity of EphB proteins. Together, our data provide biochemical and functional evidence for signal integration between Reelin and EphB forward signaling.
Figures
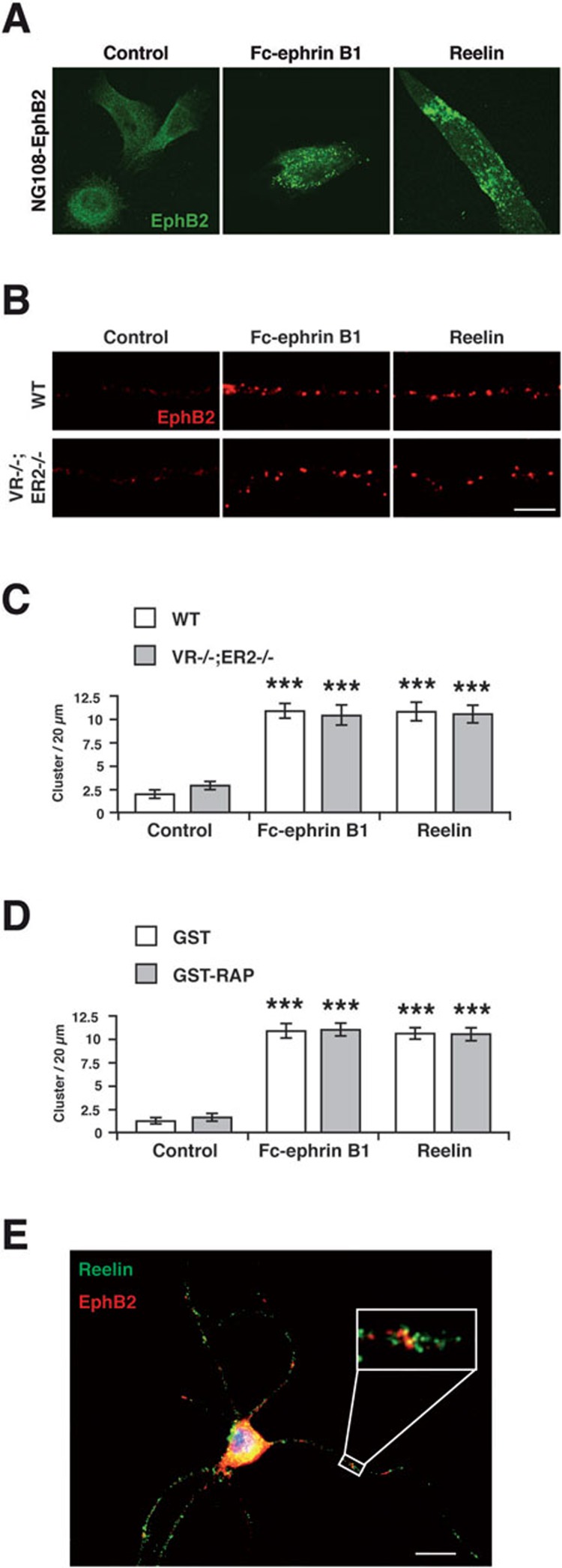
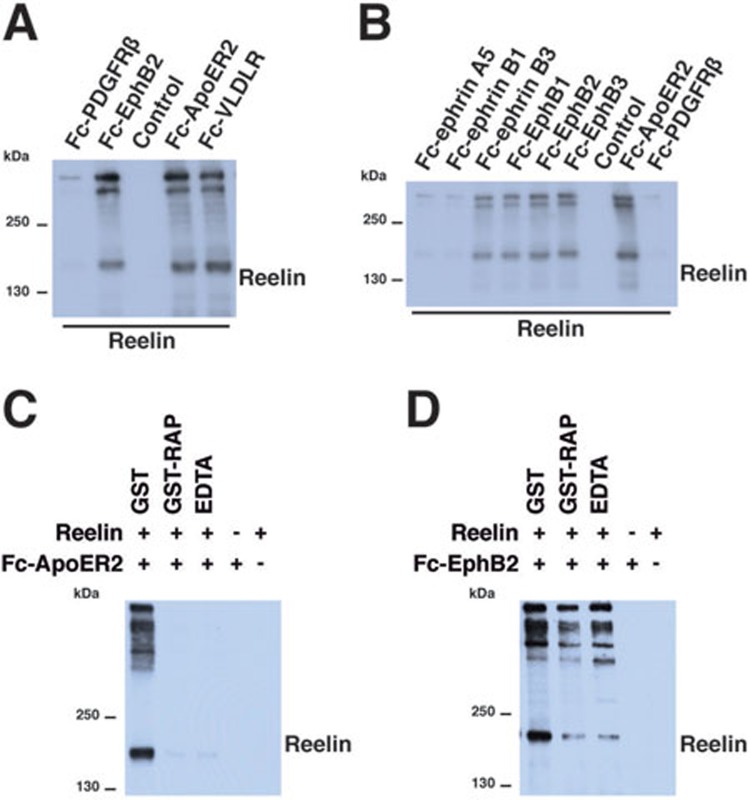
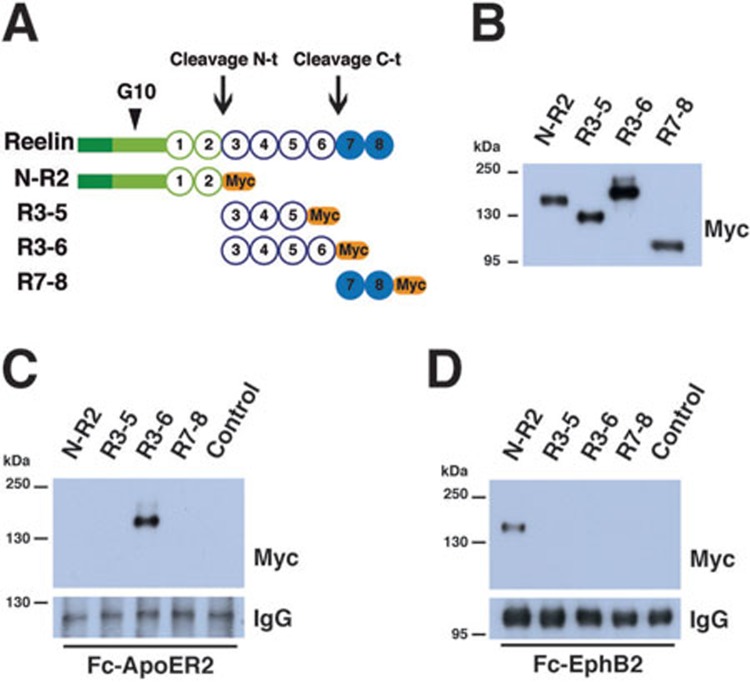
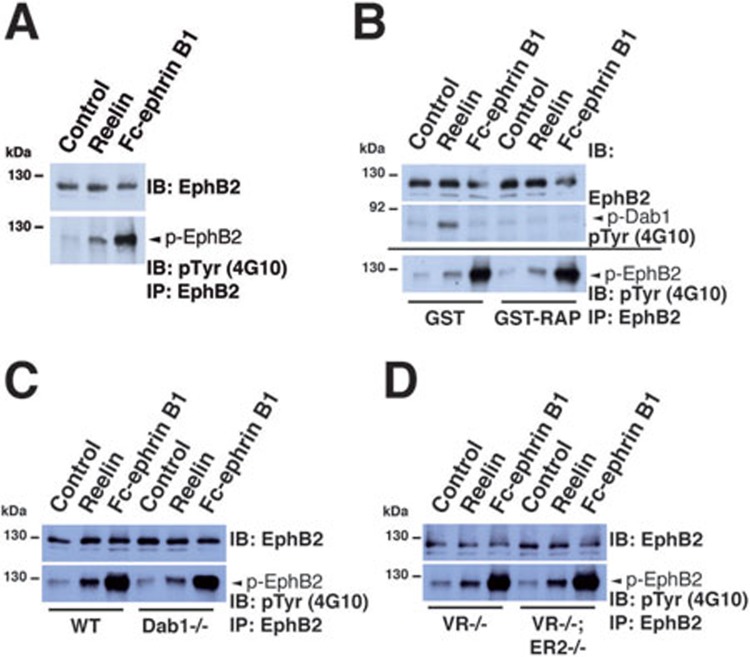
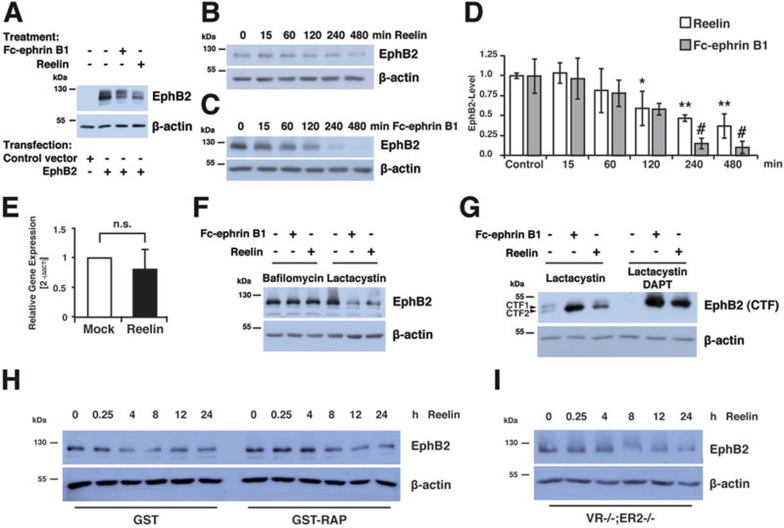
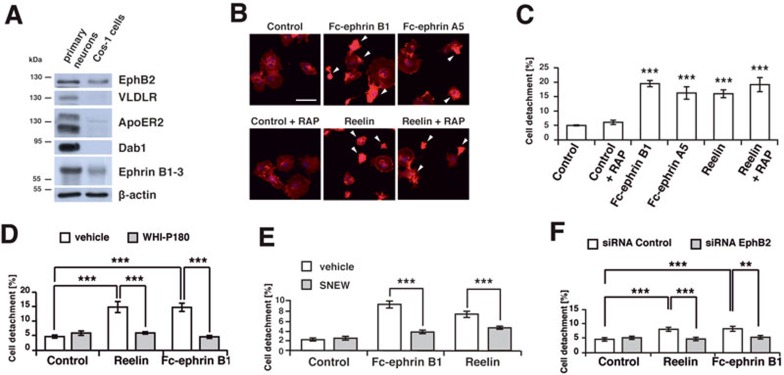
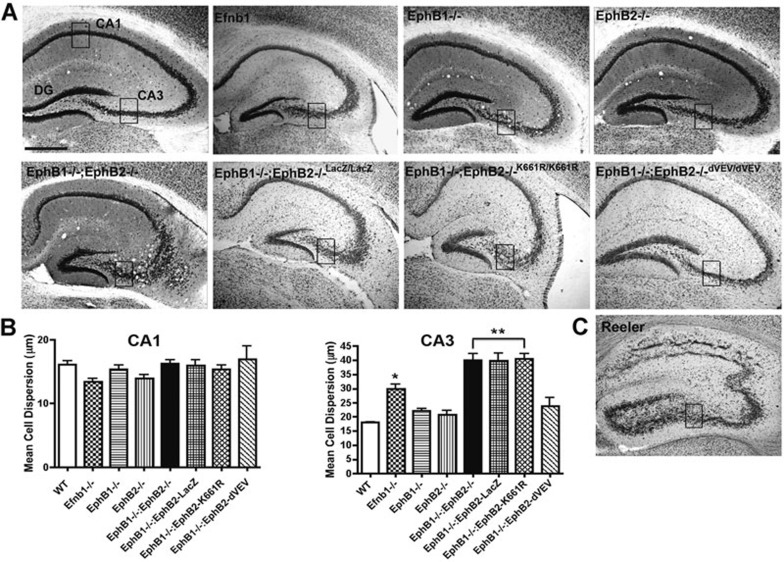
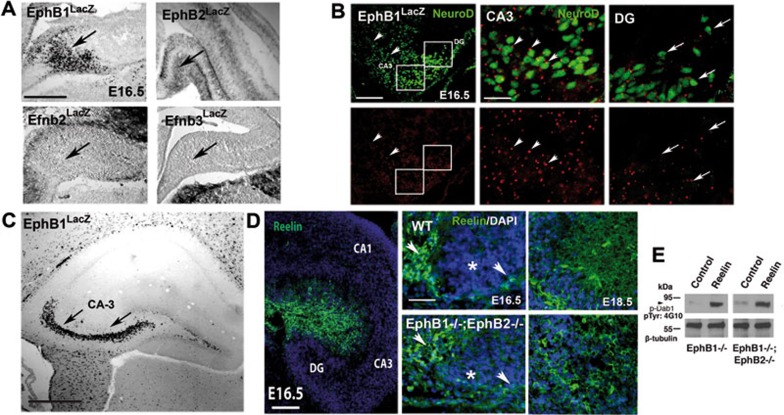
References
-
- Ayala R, Shu T, Tsai L-H. Trekking across the brain: the journey of neuronal migration. Cell. 2007;128:29–43. - PubMed
-
- Rice DS, Curran T. Role of the reelin signaling pathway in central nervous system development. Annu Rev Neurosci. 2001;24:1005–1039. - PubMed
-
- Herz J, Chen Y. Reelin, lipoprotein receptors and synaptic plasticity. Nat Rev Neurosci. 2006;7:850–859. - PubMed
-
- Tissir F, Goffinet AM. Reelin and brain development. Nat Rev Neurosci. 2003;4:496–505. - PubMed
-
- Stolt PC, Bock HH. Modulation of lipoprotein receptor functions by intracellular adaptor proteins. Cell Signal. 2006;18:1560–1571. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
