

Sequence-specific, RNA-protein interactions overcome electrostatic barriers preventing assembly of satellite tobacco necrosis virus coat protein
- PMID: 23318955
- PMCID: PMC3593212
- DOI: 10.1016/j.jmb.2013.01.004
Sequence-specific, RNA-protein interactions overcome electrostatic barriers preventing assembly of satellite tobacco necrosis virus coat protein
Abstract
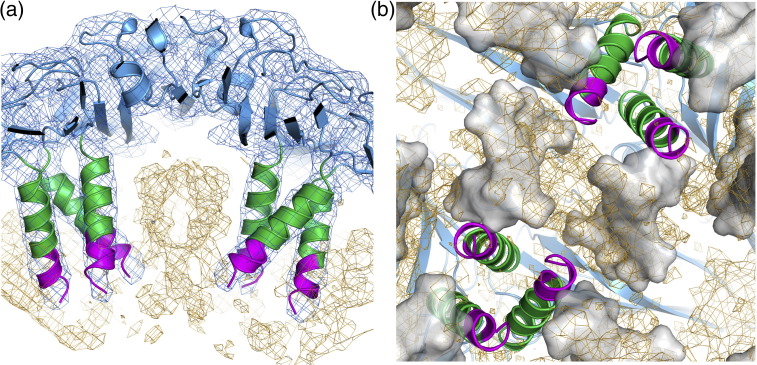
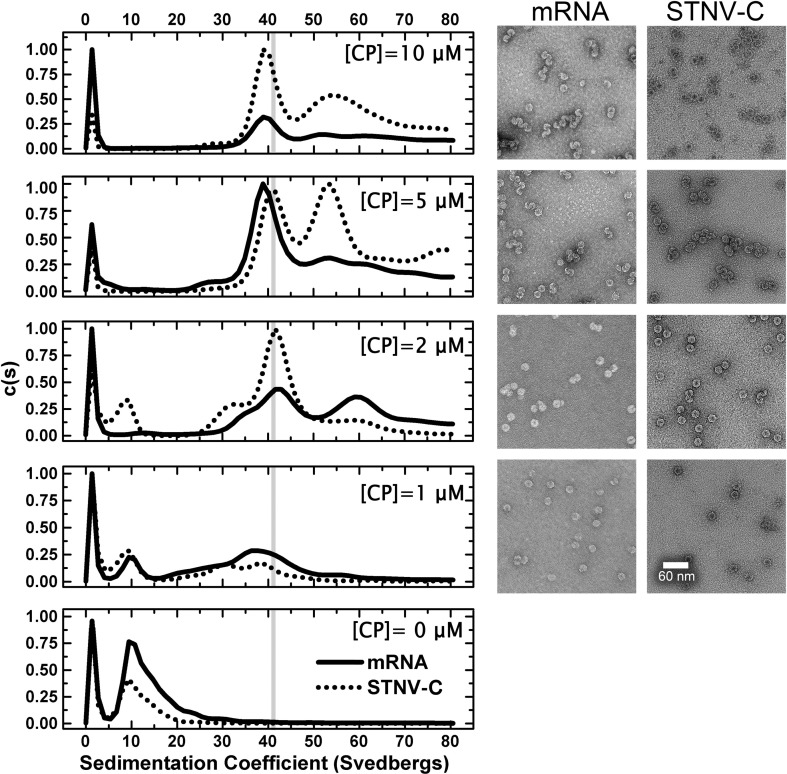
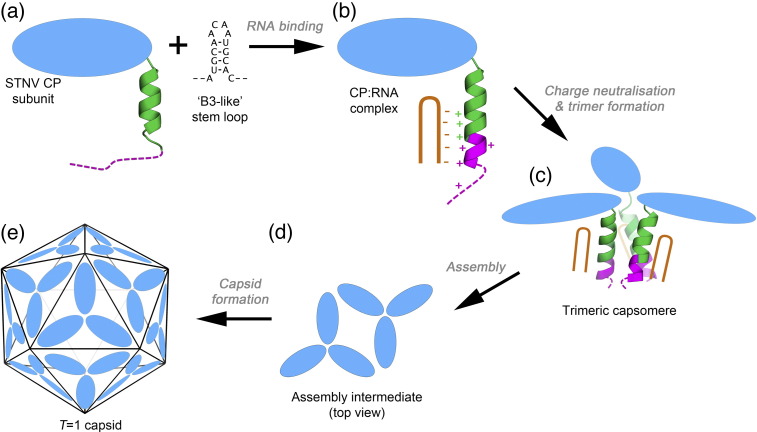
We have examined the roles of RNA-coat protein (CP) interactions in the assembly of satellite tobacco necrosis virus (STNV). The viral genomic RNA encodes only the CP, which comprises a β-barrel domain connected to a positively charged N-terminal extension. In the previous crystal structures of this system, the first 11 residues of the protein are disordered. Using variants of an RNA aptamer sequence isolated against the CP, B3, we have studied the sequence specificity of RNA-induced assembly. B3 consists of a stem-loop presenting the tetra-loop sequence ACAA. There is a clear preference for RNAs encompassing this loop sequence, as measured by the yield of T=1 capsids, which is indifferent to sequences within the stem. The B3-containing virus-like particle has been crystallised and its structure was determined to 2.3Å. A lower-resolution map encompassing density for the RNA has also been calculated. The presence of B3 results in increased ordering of the N-terminal helices located at the particle 3-fold axes, which extend by roughly one and a half turns to encompass residues 8-11, including R8 and K9. Under assembly conditions, STNV CP in the absence of RNA is monomeric and does not self-assemble. These facts suggest that a plausible model for assembly initiation is the specific RNA-induced stabilisation of a trimeric capsomere. The basic nature of the helical extension suggests that electrostatic repulsion between CPs prevents assembly in the absence of RNA and that this barrier is overcome by correct placement of appropriately orientated helical RNA stems. Such a mechanism would be consistent with the data shown here for assembly with longer RNA fragments, including an STNV genome. The results are discussed in light of a first stage of assembly involving compaction of the genomic RNA driven by multiple RNA packaging signal-CP interactions.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
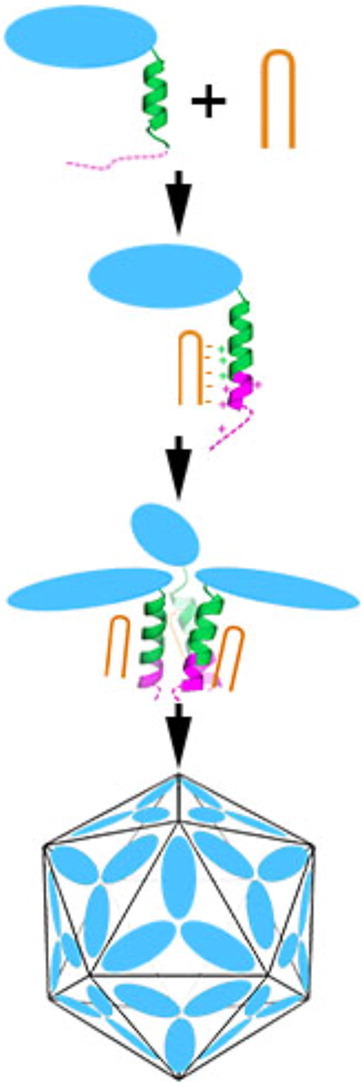
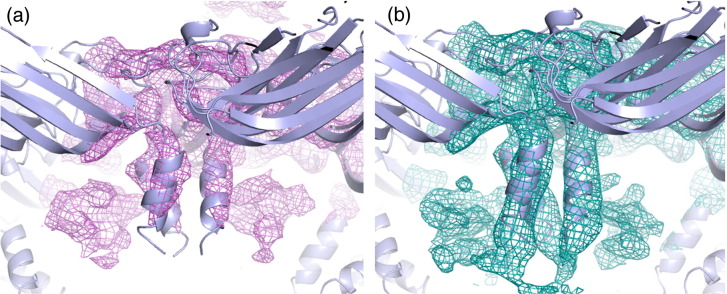
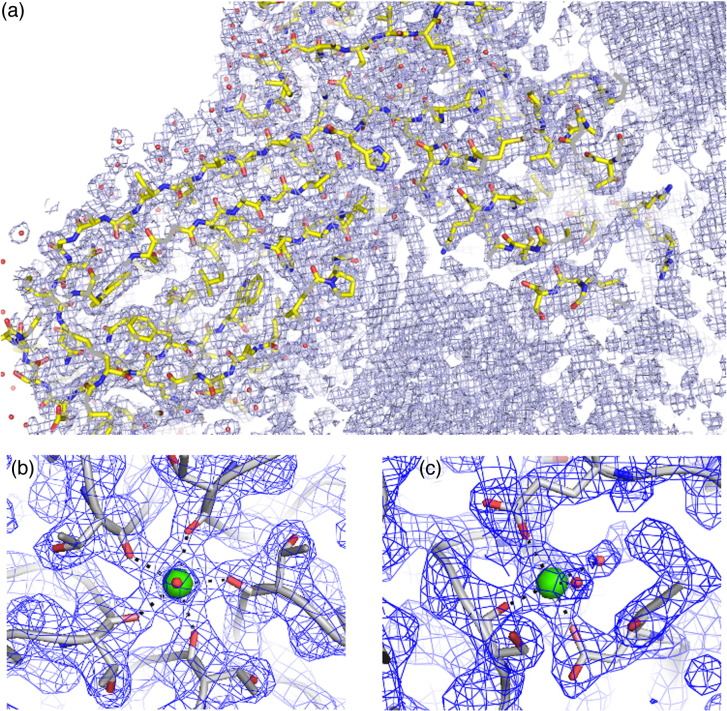
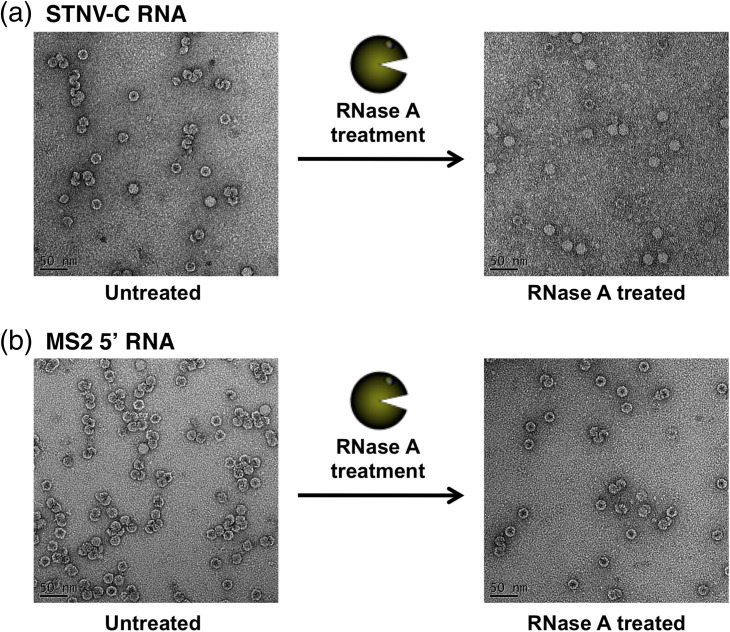
References
-
- Schneemann A. The structural and functional role of RNA in icosahedral virus assembly. Annu. Rev. Microbiol. 2006;60:51–67. - PubMed
-
- Stockley, P. G., Ranson, N. A. & Twarock, R. (2013). A new paradigm for the roles of the genome in ssRNA viruses. Future Virol. in press.
-
- Bancroft J.B., Hills G.J., Markham R. A study of the self-assembly process in a small spherical virus formation of organized structures from protein subunits in vitro. Virology. 1967;31:354–379. - PubMed
-
- Harrison S.C., Olson A.J., Schutt C.E., Winkler F.K., Bricogne G. Tomato bushy stunt virus at 2.9 Å resolution. Nature. 1978;276:368–373. - PubMed
-
- Caspar D.L.D., Klug A. Physical principles in the construction of regular viruses. Cold Spring Harbor Symp. Quant. Biol. 1962;27:1–24. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
