

Frustration in the energy landscapes of multidomain protein misfolding
- PMID: 23319605
- PMCID: PMC3562767
- DOI: 10.1073/pnas.1222130110
Frustration in the energy landscapes of multidomain protein misfolding
Abstract
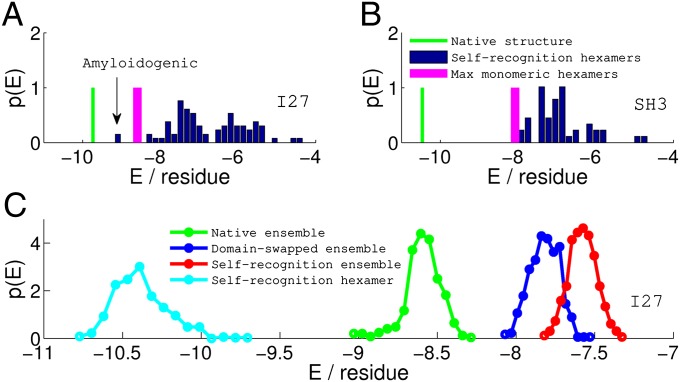
Frustration from strong interdomain interactions can make misfolding a more severe problem in multidomain proteins than in single-domain proteins. On the basis of bioinformatic surveys, it has been suggested that lowering the sequence identity between neighboring domains is one of nature's solutions to the multidomain misfolding problem. We investigate folding of multidomain proteins using the associative-memory, water-mediated, structure and energy model (AWSEM), a predictive coarse-grained protein force field. We find that reducing sequence identity not only decreases the formation of domain-swapped contacts but also decreases the formation of strong self-recognition contacts between β-strands with high hydrophobic content. The ensembles of misfolded structures that result from forming these amyloid-like interactions are energetically disfavored compared with the native state, but entropically favored. Therefore, these ensembles are more stable than the native ensemble under denaturing conditions, such as high temperature. Domain-swapped contacts compete with self-recognition contacts in forming various trapped states, and point mutations can shift the balance between the two types of interaction. We predict that multidomain proteins that lack these specific strong interdomain interactions should fold reliably.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
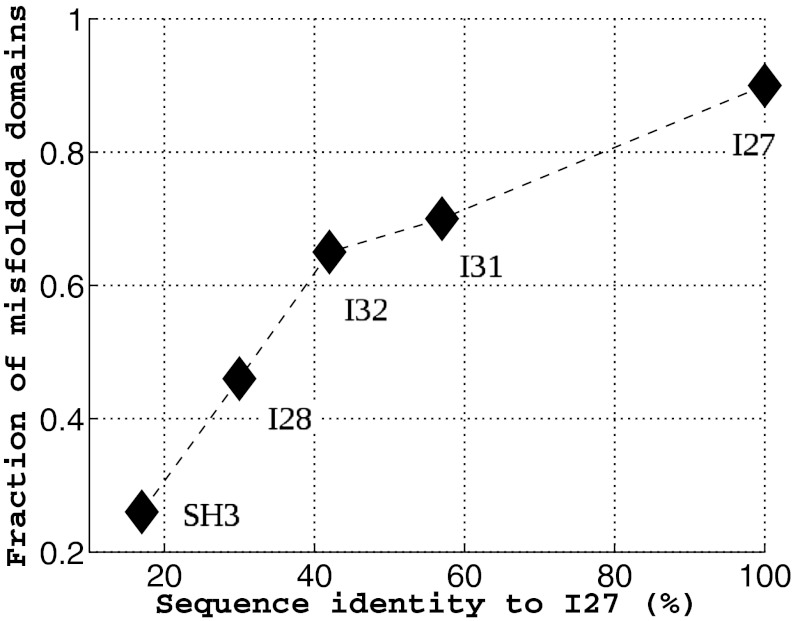
 , i = 1, 2 at the end of all of the simulations. As the sequence identity increases, the fraction of misfolded domains increases accordingly.
, i = 1, 2 at the end of all of the simulations. As the sequence identity increases, the fraction of misfolded domains increases accordingly.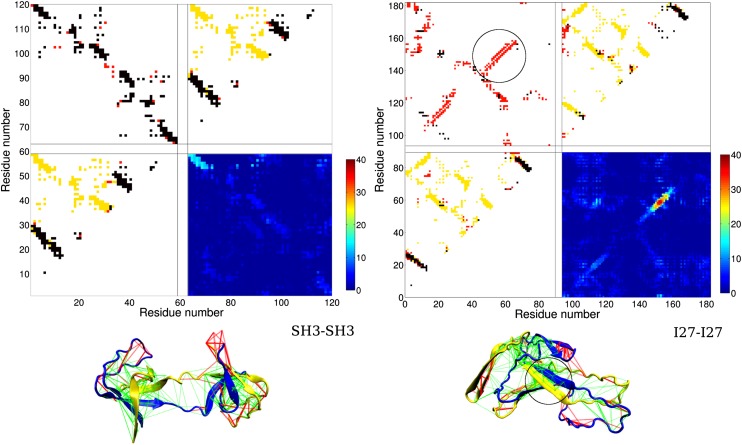
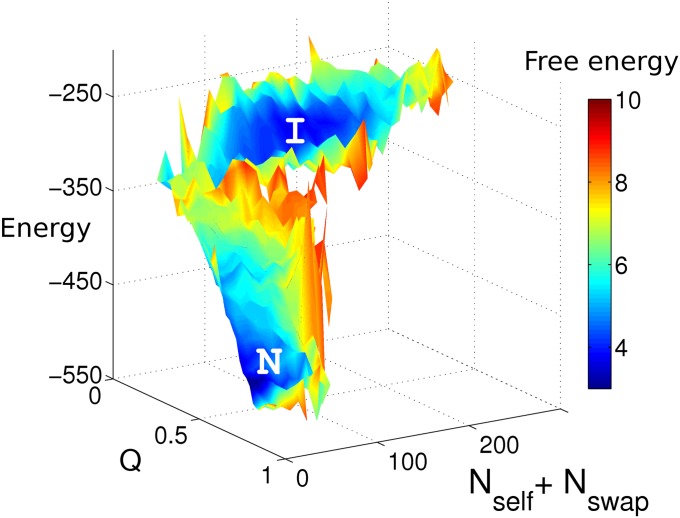
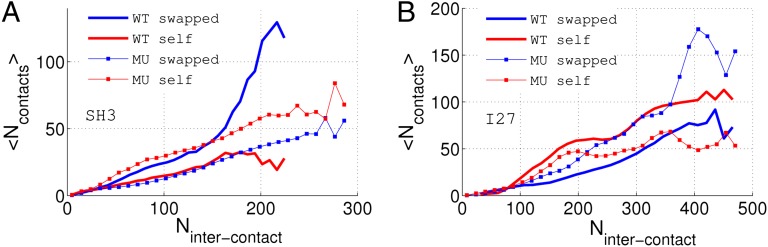
References
-
- Wolynes PG. Energy landscapes and solved protein-folding problems. Philos Trans R Soc A. 2005;363:453–467. - PubMed
-
- Selkoe DJ. Folding proteins in fatal ways. Nature. 2003;426(6968):900–904. - PubMed
-
- Chiti F, Dobson CM. Protein misfolding, functional amyloid, and human disease. Annu Rev Biochem. 2006;75:333–366. - PubMed
-
- Silow M, Tan Y-J, Fersht AR, Oliveberg M. Formation of short-lived protein aggregates directly from the coil in two-state folding. Biochemistry. 1999;38(40):13006–13012. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
