

Cortical energy demands of signaling and nonsignaling components in brain are conserved across mammalian species and activity levels
- PMID: 23319606
- PMCID: PMC3587194
- DOI: 10.1073/pnas.1214912110
Cortical energy demands of signaling and nonsignaling components in brain are conserved across mammalian species and activity levels
Abstract
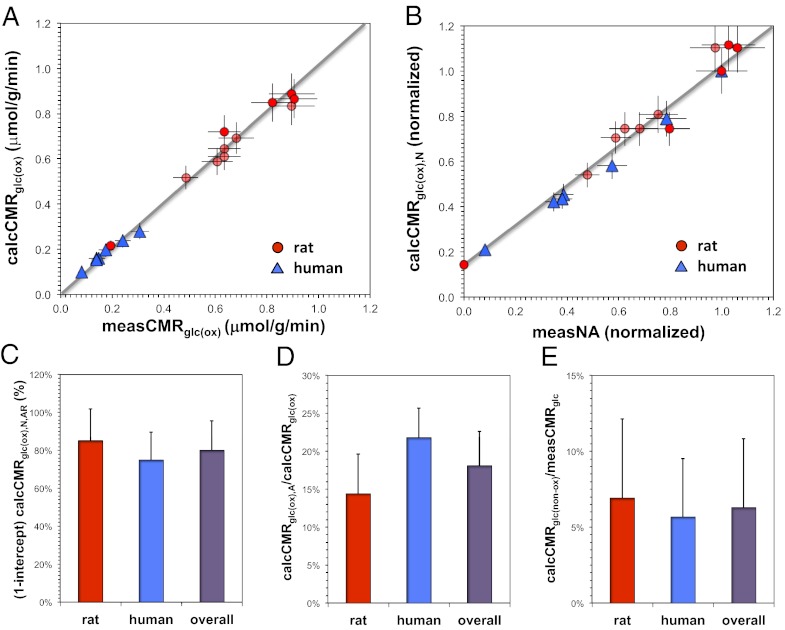
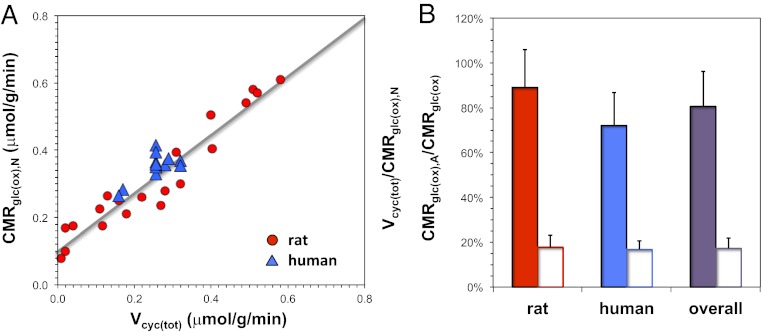
The continuous need for ion gradient restoration across the cell membrane, a prerequisite for synaptic transmission and conduction, is believed to be a major factor for brain's high oxidative demand. However, do energy requirements of signaling and nonsignaling components of cortical neurons and astrocytes vary with activity levels and across species? We derived oxidative ATP demand associated with signaling (P(s)) and nonsignaling (P(ns)) components in the cerebral cortex using species-specific physiologic and anatomic data. In rat, we calculated glucose oxidation rates from layer-specific neuronal activity measured across different states, spanning from isoelectricity to awake and sensory stimulation. We then compared these calculated glucose oxidation rates with measured glucose metabolic data for the same states as reported by 2-deoxy-glucose autoradiography. Fixed values for P(s) and P(ns) were able to predict the entire range of states in the rat. We then calculated glucose oxidation rates from human EEG data acquired under various conditions using fixed P(s) and P(ns) values derived for the rat. These calculated metabolic data in human cerebral cortex compared well with glucose metabolism measured by PET. Independent of species, linear relationship was established between neuronal activity and neuronal oxidative demand beyond isoelectricity. Cortical signaling requirements dominated energy demand in the awake state, whereas nonsignaling requirements were ∼20% of awake value. These predictions are supported by (13)C magnetic resonance spectroscopy results. We conclude that mitochondrial energy support for signaling and nonsignaling components in cerebral cortex are conserved across activity levels in mammalian species.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Allometry of brain metabolism.Proc Natl Acad Sci U S A. 2013 Feb 26;110(9):3216-7. doi: 10.1073/pnas.1221313110. Epub 2013 Feb 13. Proc Natl Acad Sci U S A. 2013. PMID: 23407170 Free PMC article. No abstract available.
References
-
- Aiello LC, Wheeler P. The expensive-tissue hypothesis—the brain and the digestive-system in human and primate evolution. Curr Anthropol. 1995;36(2):199–221.
-
- Hyder F, et al. Neuronal-glial glucose oxidation and glutamatergic-GABAergic function. J Cereb Blood Flow Metab. 2006;26(7):865–877. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
