

Cytokinins can act as suppressors of nitric oxide in Arabidopsis
- PMID: 23319631
- PMCID: PMC3557067
- DOI: 10.1073/pnas.1213235110
Cytokinins can act as suppressors of nitric oxide in Arabidopsis
Abstract
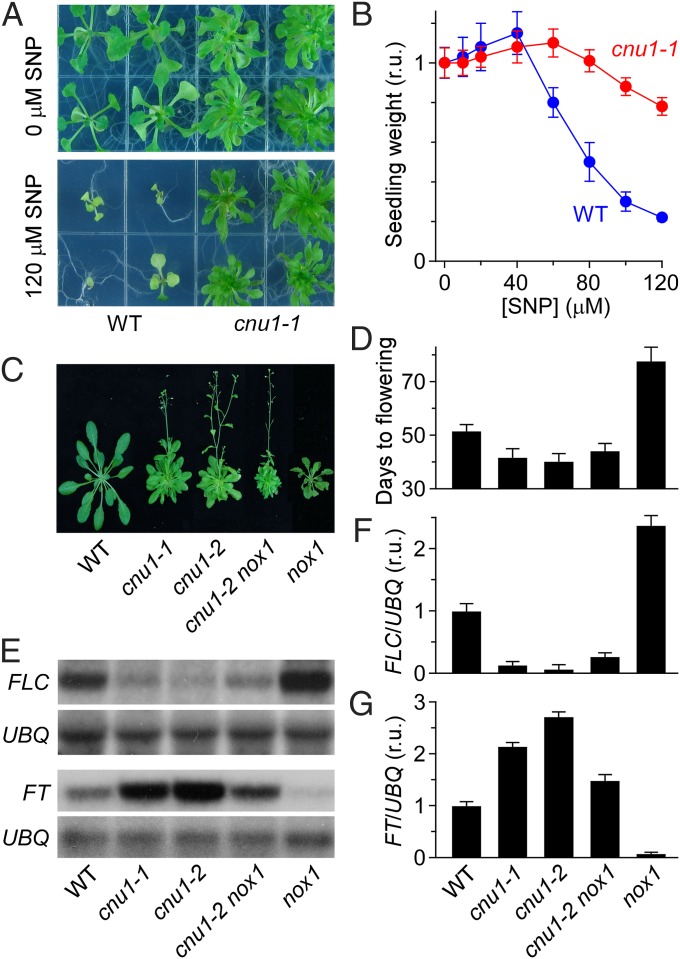
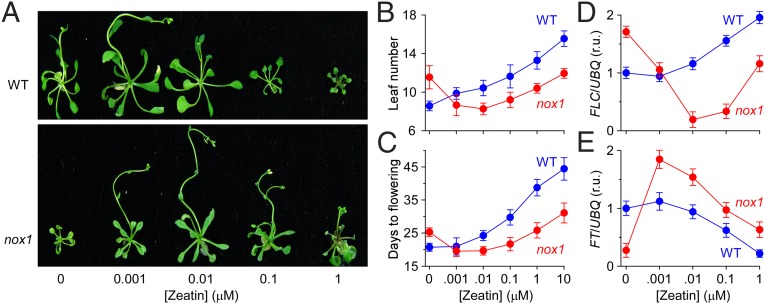
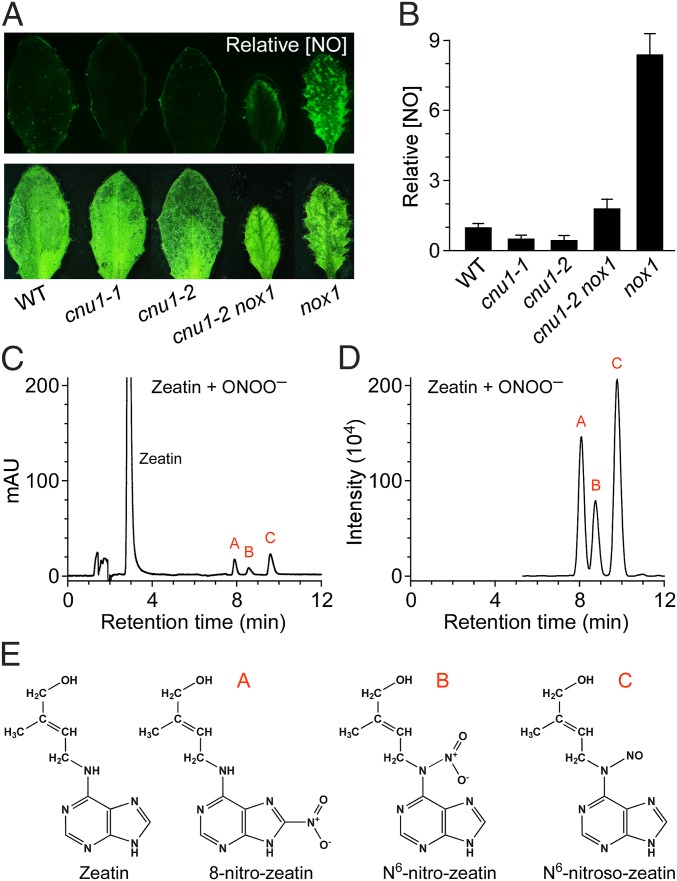
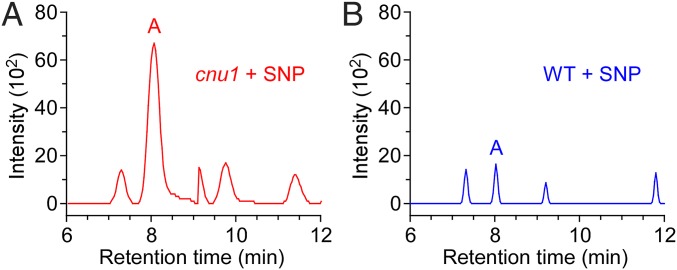
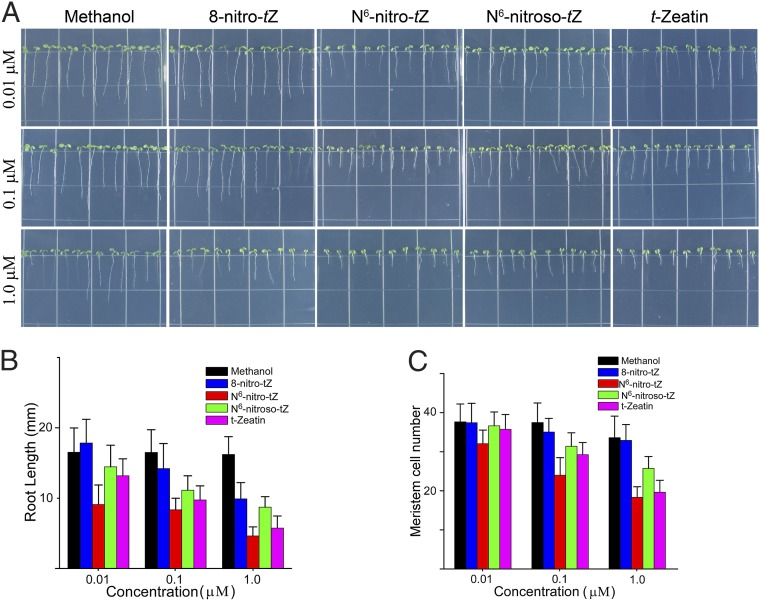
Maintaining nitric oxide (NO) homeostasis is essential for normal plant physiological processes. However, very little is known about the mechanisms of NO modulation in plants. Here, we report a unique mechanism for the catabolism of NO based on the reaction with the plant hormone cytokinin. We screened for NO-insensitive mutants in Arabidopsis and isolated two allelic lines, cnu1-1 and 1-2 (continuous NO-unstressed 1), that were identified as the previously reported altered meristem program 1 (amp1) and as having elevated levels of cytokinins. A double mutant of cnu1-2 and nitric oxide overexpression 1 (nox1) reduced the severity of the phenotypes ascribed to excess NO levels as did treating the nox1 line with trans-zeatin, the predominant form of cytokinin in Arabidopsis. We further showed that peroxinitrite, an active NO derivative, can react with zeatin in vitro, which together with the results in vivo suggests that cytokinins suppress the action of NO most likely through direct interaction between them, leading to the reduction of endogenous NO levels. These results provide insights into NO signaling and regulation of its bioactivity in plants.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Schmidt HH, Walter U. NO at work. Cell. 1994;78(6):919–925. - PubMed
-
- Lamattina L, García-Mata C, Graziano M, Pagnussat G. Nitric oxide: the versatility of an extensive signal molecule. Annu Rev Plant Biol. 2003;54:109–136. - PubMed
-
- Besson-Bard A, Pugin A, Wendehenne D. New insights into nitric oxide signaling in plants. Annu Rev Plant Biol. 2008;59:21–39. - PubMed
-
- He Y, et al. Nitric oxide represses the Arabidopsis floral transition. Science. 2004;305(5692):1968–1971. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
