

doi: 10.1085/jgp.201210888.
Epub 2013 Jan 14.
C-type inactivation of voltage-gated K+ channels: pore constriction or dilation?
Affiliations
- PMID: 23319730
- PMCID: PMC3557304
- DOI: 10.1085/jgp.201210888
Item in Clipboard
C-type inactivation of voltage-gated K+ channels: pore constriction or dilation?
J Gen Physiol.
2013 Feb.
No abstract available
Figures
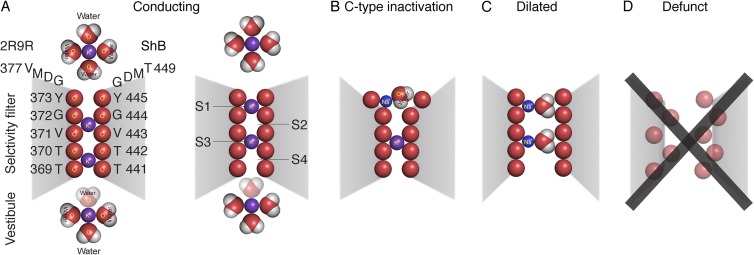
Postulated occupancy states of the selectivity filter, shown in a section through the pore. (A) Conducting states. Two K+ ions complexed with carbonyl oxygens are thought to be in the filter at any instant, in sites 2 and 4, or sites 1 and 3. Occupancy of sites 2 and 4 is more probable at resting Vm. When the gate opens, IK is normally outward, and occupancy oscillates between sites 4,2 and 3,1. This oscillation is driven by voltage, and outward movement of the ion shown in the vestibule, which sheds its watery coat and enters site 4. The ion in site 1 is driven to the site shown just external to the filter as it rehydrates. The numbering of the amino acid residues in the filter in and just above the filter of 2R9R (left) and ShB (right) are shown. (B) Postulate of a C-type inactivated channel: the outermost site 1 of the selectivity filter dilates and cannot effectively complex a K+ ion, and thus cannot accept an ion moving outward from site 2. A hydrated Na+ ion may be present in site 1, as shown. (C) Dilated state in the complete absence of K+ ions. The selectivity filter dilates from top to bottom when close-fitting K+ ions are not present and the carbonyl oxygens of the filter mutually repel. This allows partially hydrated Na+ and other large cations to permeate. (D) Defunct state. The selectivity filter is hopelessly denatured by carbonyl–carbonyl repulsion. Ion permeation is not possible. Structural details of the defunct state are unknown.
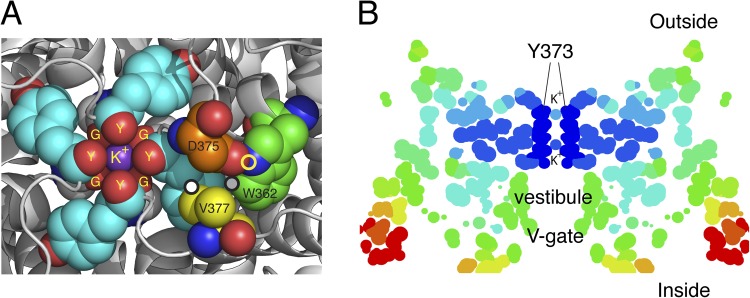
Structural arrangements of the Kv1.2-2.1 chimera (2R9R) near the pore. (A) The outermost K+ ion in site 1 (S1) of the filter is tightly complexed to the four carbonyls of Y373 (Y445) (Y) and the four carbonyls of G372 (G444) (G), just below. Several residues important in C-type inactivation are labeled. The carbon atoms of these residues are rendered with different colors for clarity: gray, G372 (Y444) in ShB; cyan, Y373 (Y445); orange, D375 (D447); yellow, V377 (T449); green, W362 (W434). Y373 (Y445) and G372 (G444) from all four subunits are shown, but only one set of D375 (D447), W362 (W434), and V377 (T449) is shown as spheres. D375/447 and W362/434 residues shown are from one subunit, whereas V377 (T449) is from an adjacent subunit. The circles mark three of the close contacts postulated to be important for C-type inactivation (see text). Prepared using MacPyMol (version 0.99). (B) A “temperature” map of a section obtained from the B-factor values along the pore axis of 2R9R. Dark blue marks regions of high stability, ranging through light blue, yellow, orange, and red as stability decreases progressively away from the selectivity filter. Prepared using RasMol.
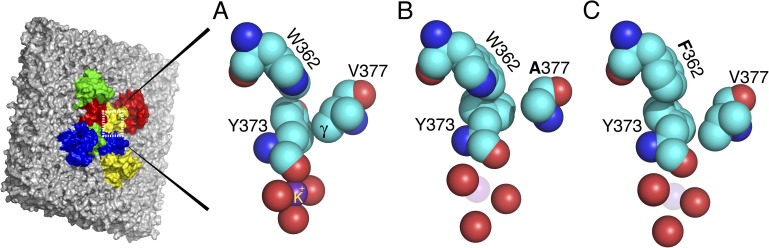
An expanded view of the outer mouth of the selectivity filter of the Kv1.2-2.1 chimera and two simulated mutations. The four outermost Y373 carbonyl oxygens outline the pore, but only one tyrosine is shown in full. (A) The conducting conformation of the filter mouth, taken from the Kv1.2-2.1 chimera. Y373 (Y445) has close contacts with W362 (W434) and V377 (T449). (B) A simulated mutation of the Kv1.2-2.1 chimera that replaces V377 (T449) with V377A (T449A), allowing Y373 (Y445) to rotate and dilate the filter mouth. The mutation in effect deletes the two γ carbons of V377 (T449), removing a contact that locks Y373 (Y445) in the conducting, nondilated position. (C) Simulated mutation W362 (W434) of the Kv1.2-2.1 chimera that replaces W362 (W434) with F362 (F434). Relative to W362 (W434), F362 (F434) is rotated clockwise in this view; it presses against the ring of Y373 (Y445), causing Y373 (Y445) to rotate counterclockwise, dilating the filter mouth. The angle of view is the same throughout the figure. The images were prepared using MacPyMol (version 0.99).
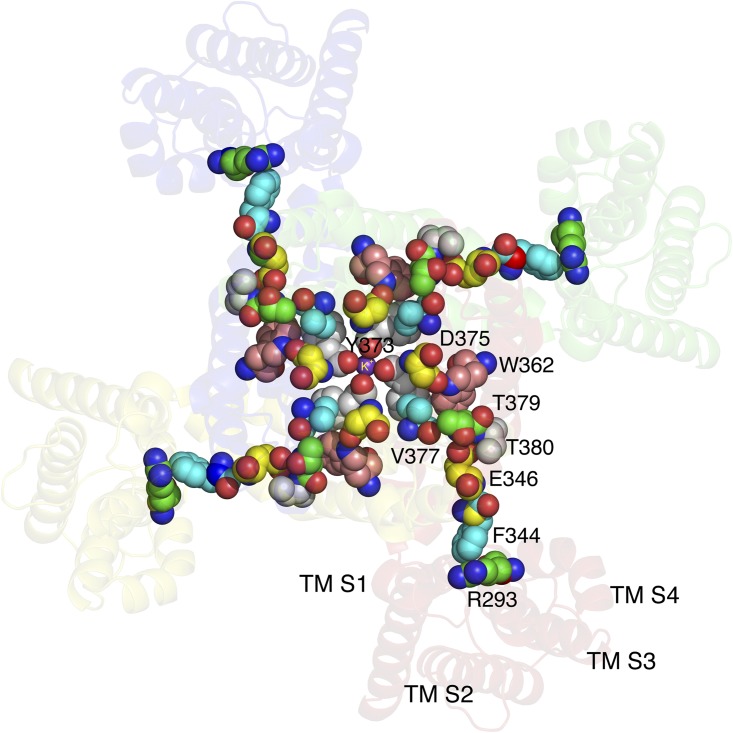
“Four-legged runner” model of the coupling of S4 activation and C-type inactivation. A Kv1.2-2.1 chimera (2R9R) is viewed from the extracellular side. Consistent with the literature, a conformational wave begins with contact between F344 in TMS5 and R293 in TMS4, when the latter is driven outward by voltage. The wave spreads through the illustrated residues to the region of the pore mouth, causing, in susceptible mutants, dilation of the pore mouth and C-type inactivation. The Kv1.2-2.1 chimera does not undergo C-type inactivation because V377 locks Y373 in position; the pore mouth here is not dilated. All of the residues presented as spheres have been shown to affect C-type inactivation. The carbon atoms of these residues are shown using different colors for clarity. TMS1–TMS4 identify the respective transmembrane segments. Prepared using MacPyMol (version 0.99).
Similar articles
-
A structural interpretation of voltage-gated potassium channel inactivation.Prog Biophys Mol Biol. 2006 Oct;92(2):185-208. doi: 10.1016/j.pbiomolbio.2005.10.001. Epub 2005 Nov 8. Prog Biophys Mol Biol. 2006. PMID: 16316679 Review.
-
Constitutive inactivation of the hKv1.5 mutant channel, H463G, in K+-free solutions at physiological pH.Cell Biochem Biophys. 2005;43(2):221-30. doi: 10.1385/CBB:43:2:221. Cell Biochem Biophys. 2005. PMID: 16049347
-
Voltage-gated ion channels.Curr Biol. 2005 Jan 26;15(2):R44-7. doi: 10.1016/j.cub.2004.12.050. Curr Biol. 2005. PMID: 15668152 Review. No abstract available.
-
The HERG K+ channel: progress in understanding the molecular basis of its unusual gating kinetics.Eur Biophys J. 2004 Apr;33(2):89-97. doi: 10.1007/s00249-003-0338-3. Epub 2003 Sep 10. Eur Biophys J. 2004. PMID: 13680209 Review.
-
Models of the structure and voltage-gating mechanism of the shaker K+ channel.Biophys J. 2004 Oct;87(4):2116-30. doi: 10.1529/biophysj.104.040618. Biophys J. 2004. PMID: 15454416 Free PMC article.
Cited by
-
Exploring the Dynamics of the TWIK-1 Channel.Biophys J. 2016 Aug 23;111(4):775-784. doi: 10.1016/j.bpj.2016.07.009. Biophys J. 2016. PMID: 27558721 Free PMC article.
-
Pore helices play a dynamic role as integrators of domain motion during Kv11.1 channel inactivation gating.J Biol Chem. 2013 Apr 19;288(16):11482-91. doi: 10.1074/jbc.M113.461442. Epub 2013 Mar 7. J Biol Chem. 2013. PMID: 23471968 Free PMC article.
-
Marine Toxins Targeting Kv1 Channels: Pharmacological Tools and Therapeutic Scaffolds.Mar Drugs. 2020 Mar 20;18(3):173. doi: 10.3390/md18030173. Mar Drugs. 2020. PMID: 32245015 Free PMC article. Review.
-
A perspective on Na and K channel inactivation.J Gen Physiol. 2018 Jan 2;150(1):7-18. doi: 10.1085/jgp.201711835. Epub 2017 Dec 12. J Gen Physiol. 2018. PMID: 29233885 Free PMC article. Review.
-
Molecular basis of PIP2-dependent regulation of the Ca2+-activated chloride channel TMEM16A.Nat Commun. 2019 Aug 21;10(1):3769. doi: 10.1038/s41467-019-11784-8. Nat Commun. 2019. PMID: 31434906 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
