

Activation of the cellular unfolded protein response by recombinant adeno-associated virus vectors
- PMID: 23320106
- PMCID: PMC3540029
- DOI: 10.1371/journal.pone.0053845
Activation of the cellular unfolded protein response by recombinant adeno-associated virus vectors
Abstract
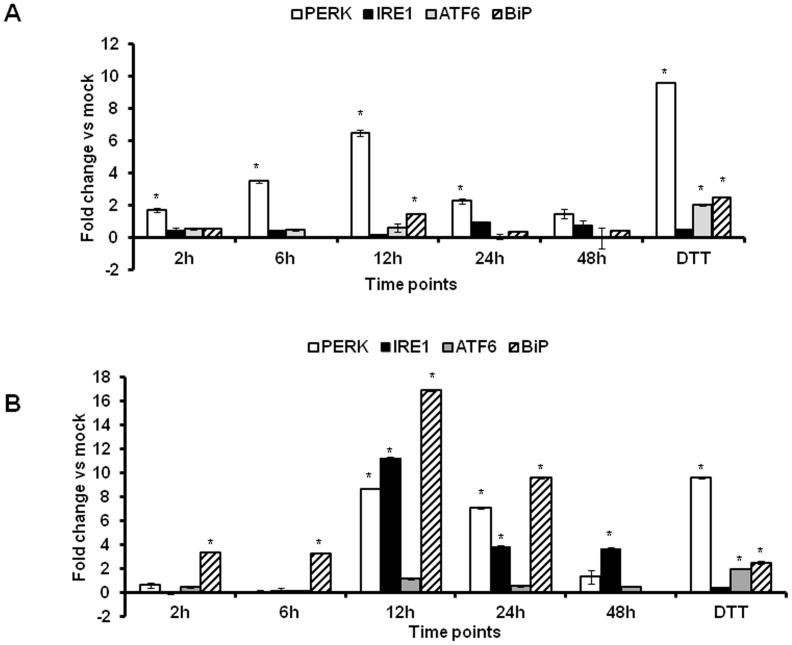
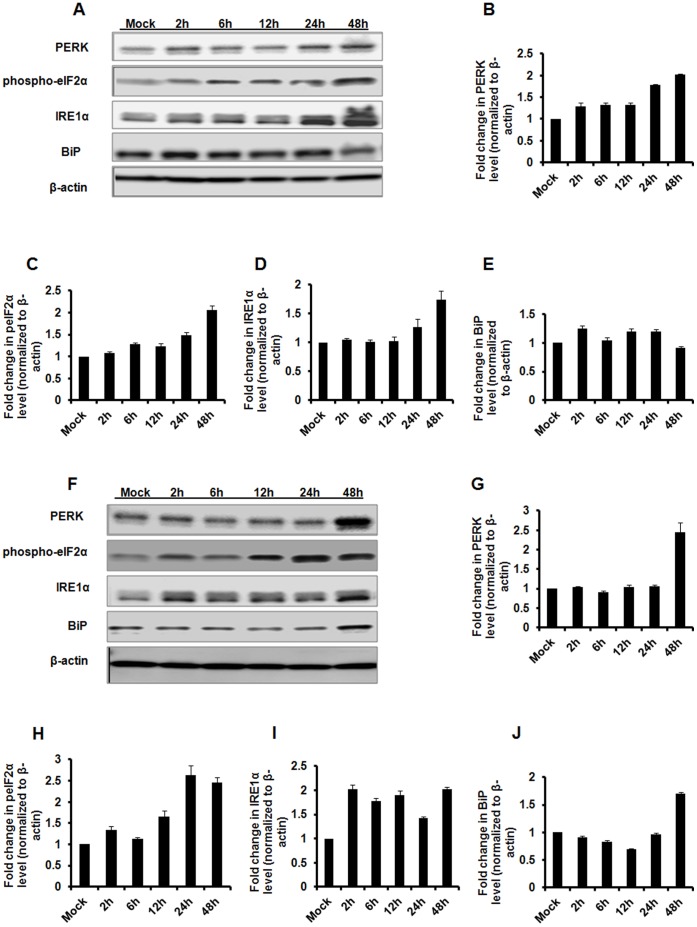
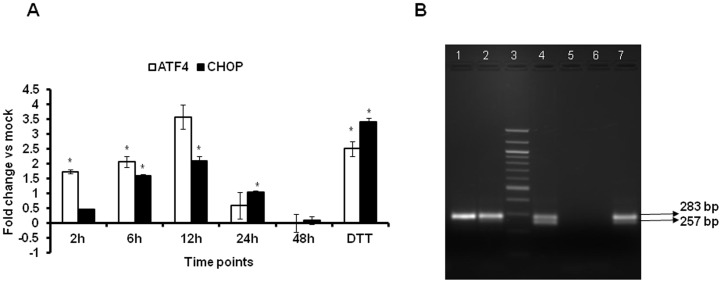
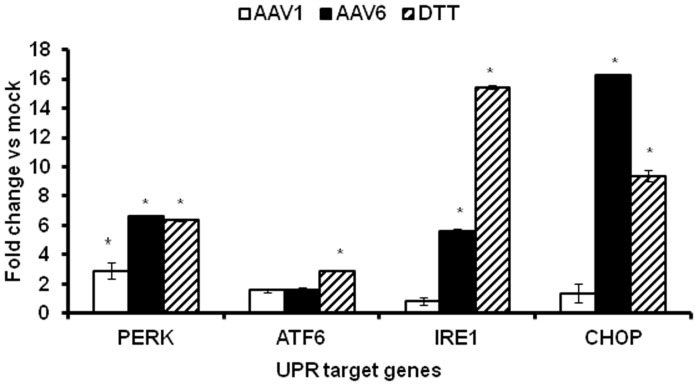
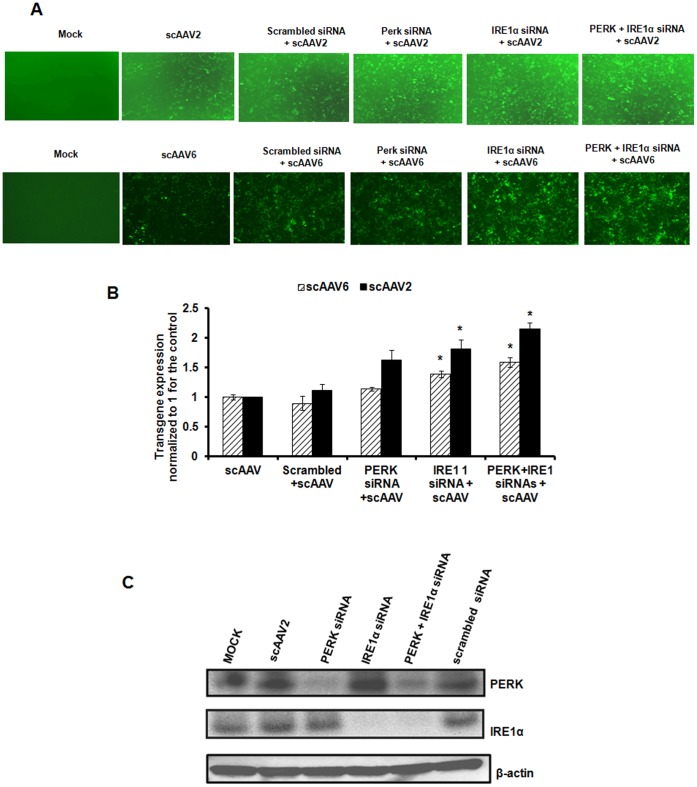
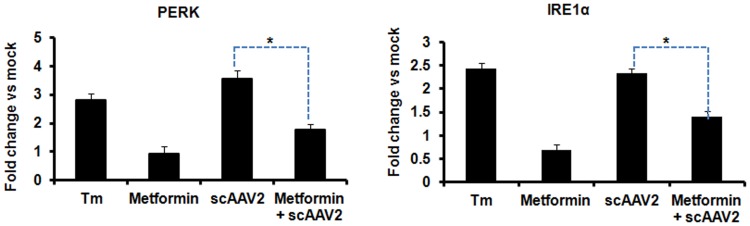
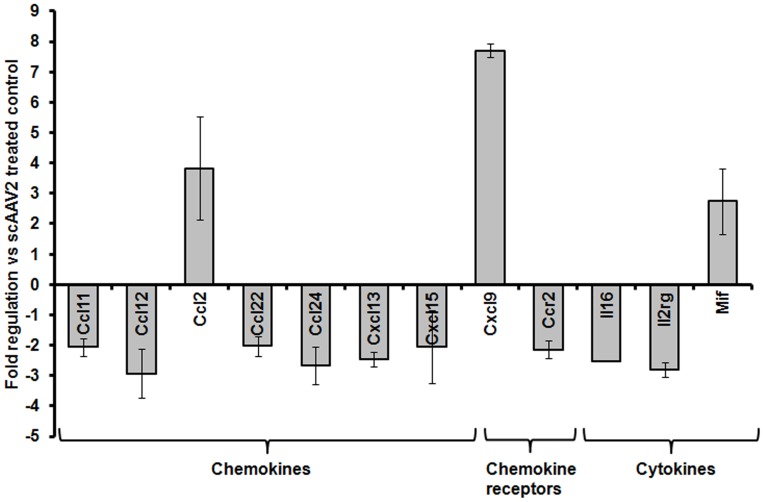
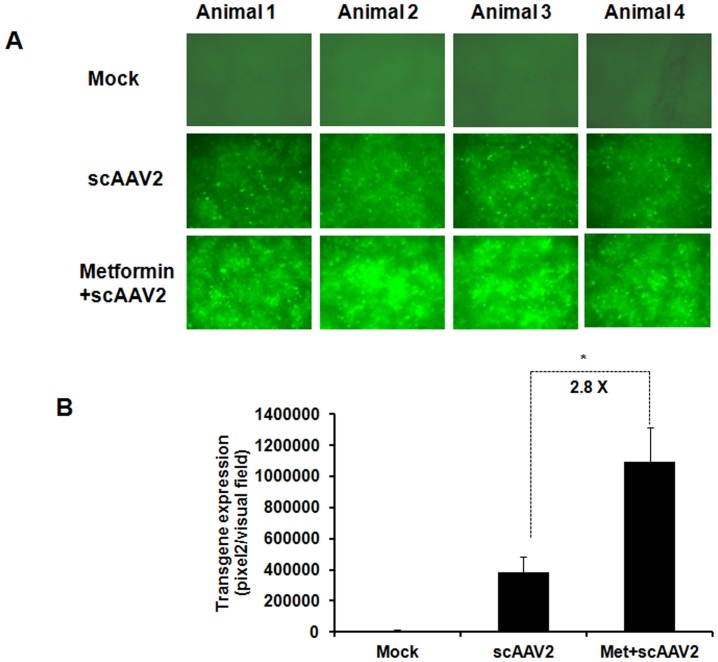
The unfolded protein response (UPR) is a stress-induced cyto-protective mechanism elicited towards an influx of large amount of proteins in the endoplasmic reticulum (ER). In the present study, we evaluated if AAV manipulates the UPR pathways during its infection. We first examined the role of the three major UPR axes, namely, endoribonuclease inositol-requiring enzyme-1 (IRE1α), activating transcription factor 6 (ATF6) and PKR-like ER kinase (PERK) in AAV infected cells. Total RNA from mock or AAV infected HeLa cells were used to determine the levels of 8 different ER-stress responsive transcripts from these pathways. We observed a significant up-regulation of IRE1α (up to 11 fold) and PERK (up to 8 fold) genes 12-48 hours after infection with self-complementary (sc)AAV2 but less prominent with single-stranded (ss)AAV2 vectors. Further studies demonstrated that scAAV1 and scAAV6 also induce cellular UPR in vitro, with AAV1 vectors activating the PERK pathway (3 fold) while AAV6 vectors induced a significant increase on all the three major UPR pathways [6-16 fold]. These data suggest that the type and strength of UPR activation is dependent on the viral capsid. We then examined if transient inhibition of UPR pathways by RNA interference has an effect on AAV transduction. siRNA mediated silencing of PERK and IRE1α had a modest effect on AAV2 and AAV6 mediated gene expression (∼1.5-2 fold) in vitro. Furthermore, hepatic gene transfer of scAAV2 vectors in vivo, strongly elevated IRE1α and PERK pathways (2 and 3.5 fold, respectively). However, when animals were pre-treated with a pharmacological UPR inhibitor (metformin) during scAAV2 gene transfer, the UPR signalling and its subsequent inflammatory response was attenuated concomitant to a modest 2.8 fold increase in transgene expression. Collectively, these data suggest that AAV vectors activate the cellular UPR pathways and their selective inhibition may be beneficial during AAV mediated gene transfer.
Conflict of interest statement
Figures
Similar articles
-
ATF6-Mediated Unfolded Protein Response Facilitates Adeno-associated Virus 2 (AAV2) Transduction by Releasing the Suppression of the AAV Receptor on Endoplasmic Reticulum Stress.J Virol. 2022 Feb 9;96(3):e0110321. doi: 10.1128/JVI.01103-21. Epub 2021 Dec 1. J Virol. 2022. PMID: 34851146 Free PMC article.
-
The unfolded protein response is triggered following a single, unaccustomed resistance-exercise bout.Am J Physiol Regul Integr Comp Physiol. 2014 Sep 15;307(6):R664-9. doi: 10.1152/ajpregu.00511.2013. Epub 2014 Jul 9. Am J Physiol Regul Integr Comp Physiol. 2014. PMID: 25009220 Clinical Trial.
-
Mechanism of the induction of endoplasmic reticulum stress by the anti-cancer agent, di-2-pyridylketone 4,4-dimethyl-3-thiosemicarbazone (Dp44mT): Activation of PERK/eIF2α, IRE1α, ATF6 and calmodulin kinase.Biochem Pharmacol. 2016 Jun 1;109:27-47. doi: 10.1016/j.bcp.2016.04.001. Epub 2016 Apr 6. Biochem Pharmacol. 2016. PMID: 27059255
-
Targeting UPR branches, a potential strategy for enhancing efficacy of cancer chemotherapy.Acta Biochim Biophys Sin (Shanghai). 2021 Nov 10;53(11):1417-1427. doi: 10.1093/abbs/gmab131. Acta Biochim Biophys Sin (Shanghai). 2021. PMID: 34664059 Review.
-
Unfolding anti-tumor immunity: ER stress responses sculpt tolerogenic myeloid cells in cancer.J Immunother Cancer. 2017 Jan 17;5:5. doi: 10.1186/s40425-016-0203-4. eCollection 2017. J Immunother Cancer. 2017. PMID: 28105371 Free PMC article. Review.
Cited by
-
Evidence against a contribution of the CCAAT-enhancer binding protein homologous protein (CHOP) in mediating neurotoxicity in rTg4510 mice.Sci Rep. 2022 May 5;12(1):7372. doi: 10.1038/s41598-022-11025-x. Sci Rep. 2022. PMID: 35513476 Free PMC article.
-
Adeno-associated virus vectors and neurotoxicity-lessons from preclinical and human studies.Gene Ther. 2025 Jan;32(1):60-73. doi: 10.1038/s41434-023-00405-1. Epub 2023 May 10. Gene Ther. 2025. PMID: 37165032 Free PMC article. Review.
-
Transcriptomics-informed pharmacology identifies epigenetic and cell cycle regulators that enhance AAV production.Mol Ther Methods Clin Dev. 2024 Nov 18;32(4):101384. doi: 10.1016/j.omtm.2024.101384. eCollection 2024 Dec 12. Mol Ther Methods Clin Dev. 2024. PMID: 39687728 Free PMC article.
-
Temporal insights into molecular and cellular responses during rAAV production in HEK293T cells.Mol Ther Methods Clin Dev. 2024 Jun 8;32(3):101278. doi: 10.1016/j.omtm.2024.101278. eCollection 2024 Sep 12. Mol Ther Methods Clin Dev. 2024. PMID: 39022743 Free PMC article.
-
Viral Infections and Their Ability to Modulate Endoplasmic Reticulum Stress Response Pathways.Viruses. 2024 Sep 30;16(10):1555. doi: 10.3390/v16101555. Viruses. 2024. PMID: 39459886 Free PMC article. Review.
References
-
- Erles K, Sebokova P, Schlehofer JR (1999) Update on the prevalence of serum antibodies (IgG and IgM) to adeno-associated virus (AAV). J Med Virol 59: 406–411. - PubMed
-
- Berns KI, Linden RM (1995) The cryptic life style of adeno-associated virus. Bioessays 17: 237–245. - PubMed
-
- Manno CS, Pierce GF, Arruda VR, Glader B, Ragni M, et al. (2006) Successful transduction of liver in hemophilia by AAV-Factor IX and limitations imposed by the host immune response. Nat Med 12: 342–347. - PubMed
-
- Mingozzi F, Maus MV, Hui DJ, Sabatino DE, Murphy SL, et al. (2007) CD8(+) T-cell responses to adeno-associated virus capsid in humans. Nat Med 13: 419–422. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
