

Katanin localization requires triplet microtubules in Chlamydomonas reinhardtii
- PMID: 23320108
- PMCID: PMC3540033
- DOI: 10.1371/journal.pone.0053940
Katanin localization requires triplet microtubules in Chlamydomonas reinhardtii
Erratum in
-
Correction: Katanin Localization Requires Triplet Microtubules in Chlamydomonas reinhardtii.PLoS One. 2016 Jan 15;11(1):e0145569. doi: 10.1371/journal.pone.0145569. eCollection 2016. PLoS One. 2016. PMID: 26771461 Free PMC article. No abstract available.
Abstract
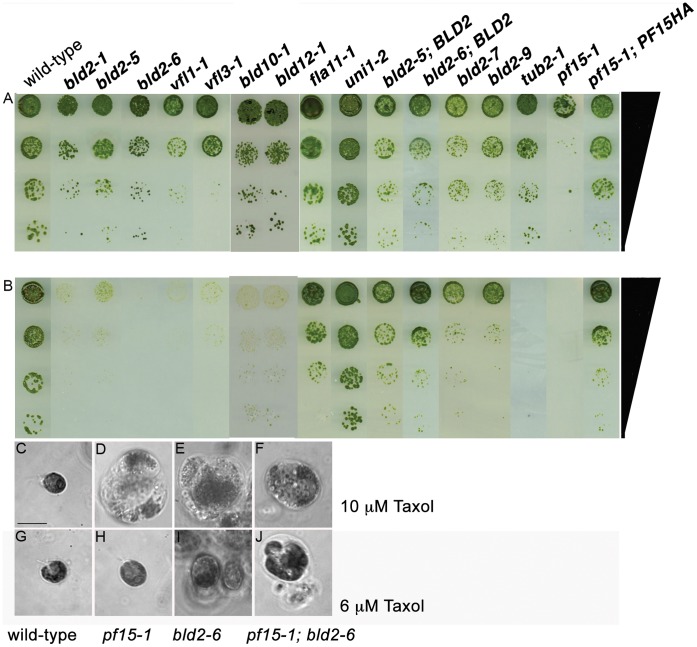
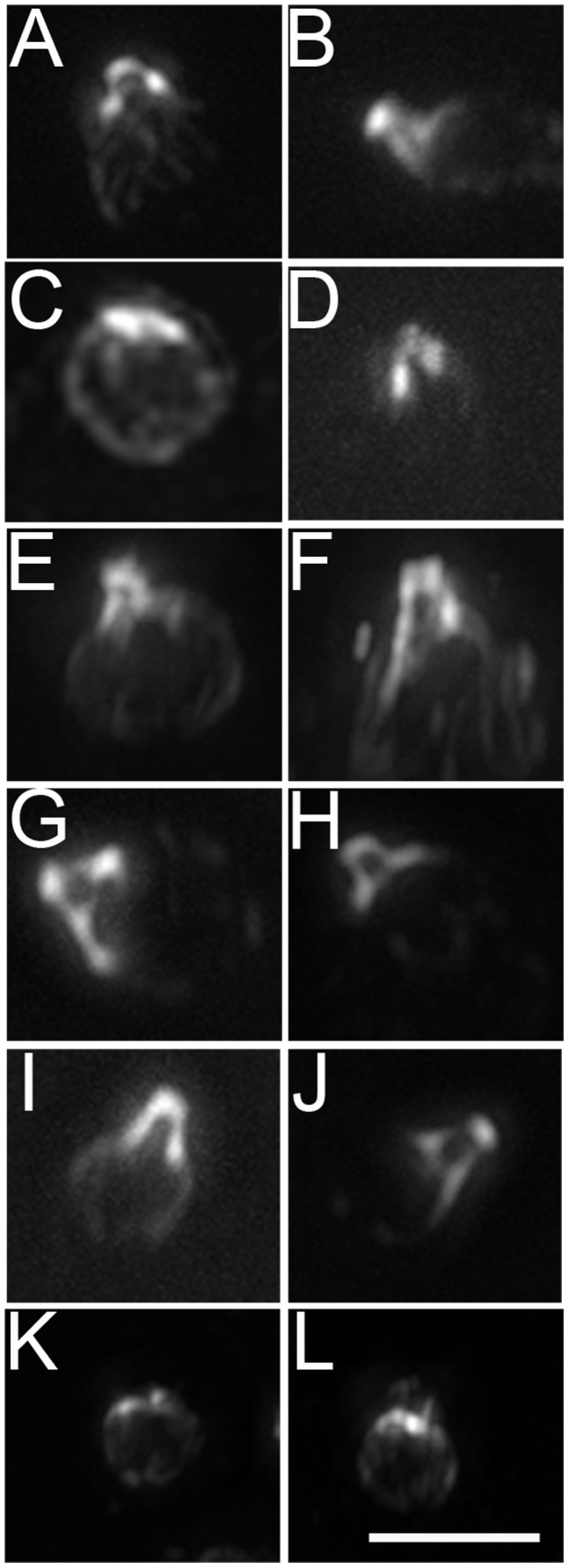
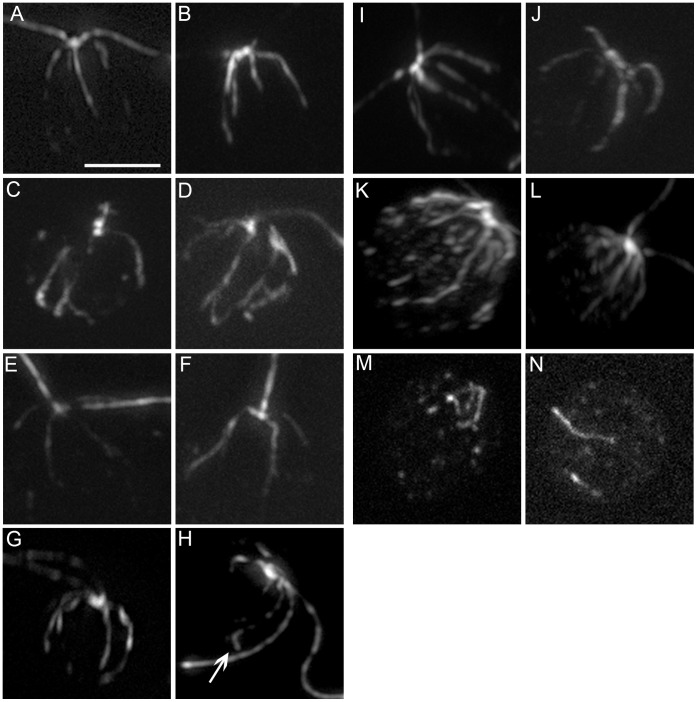
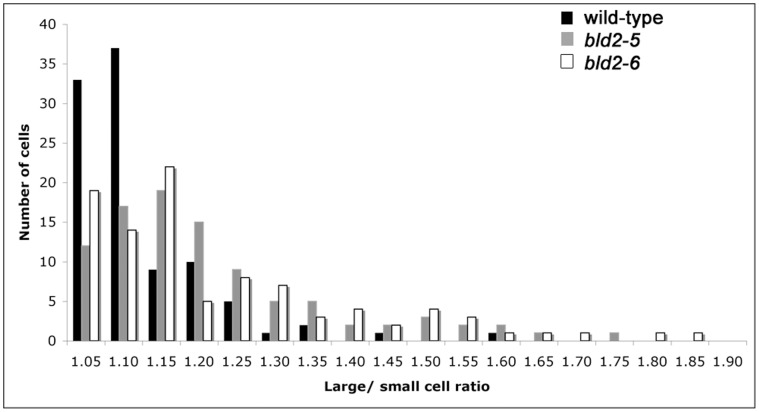
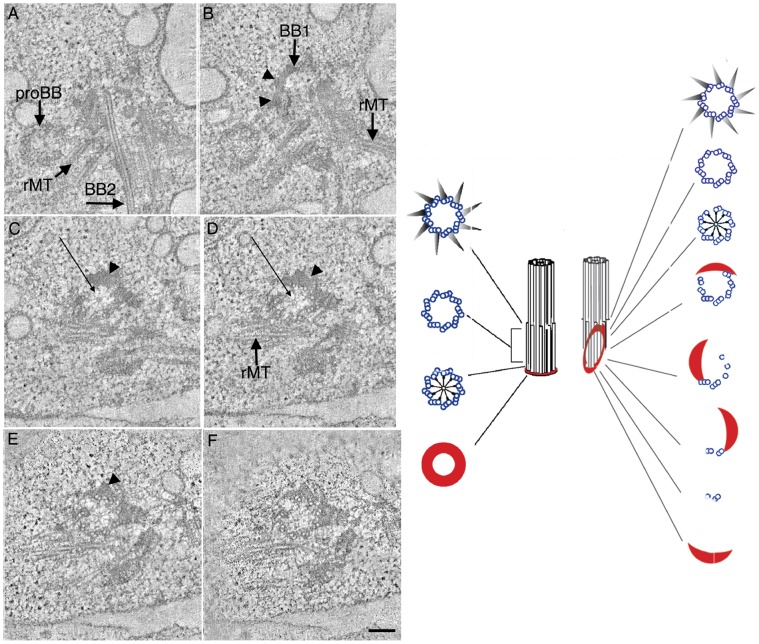
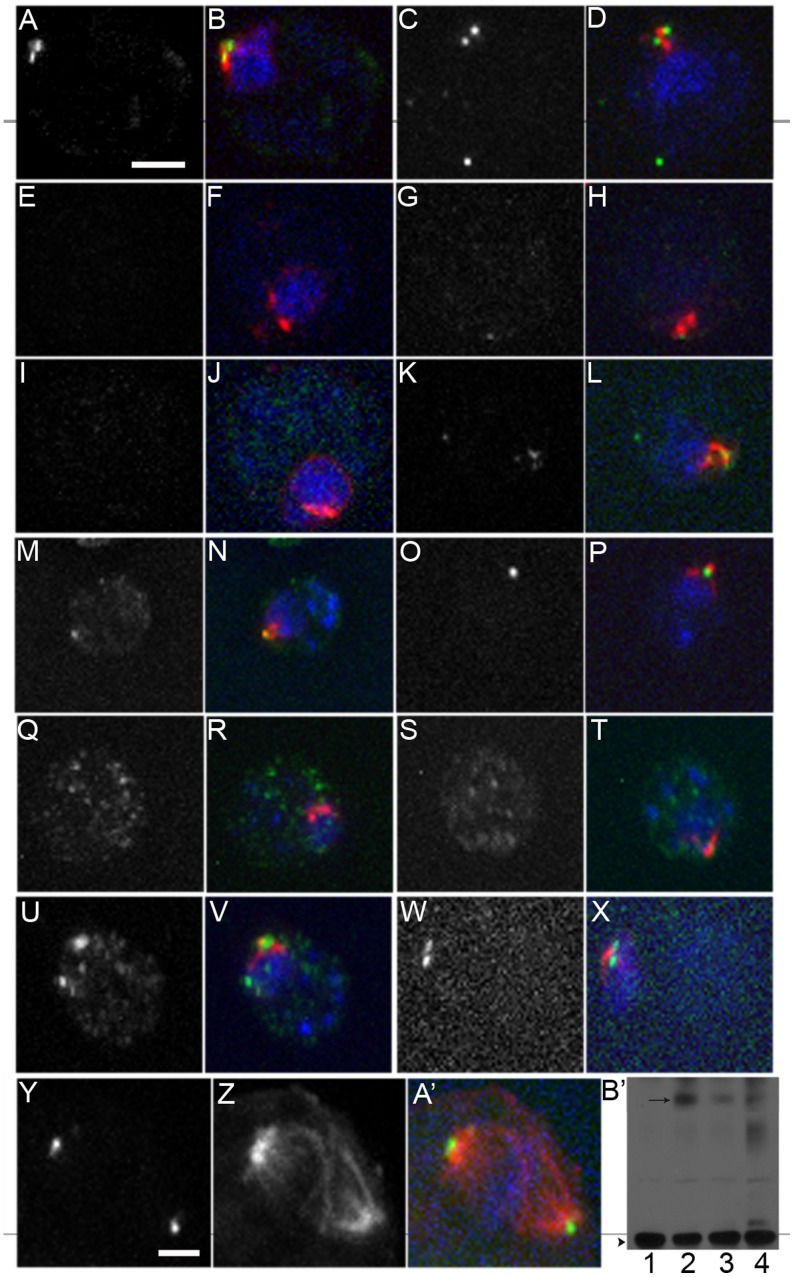
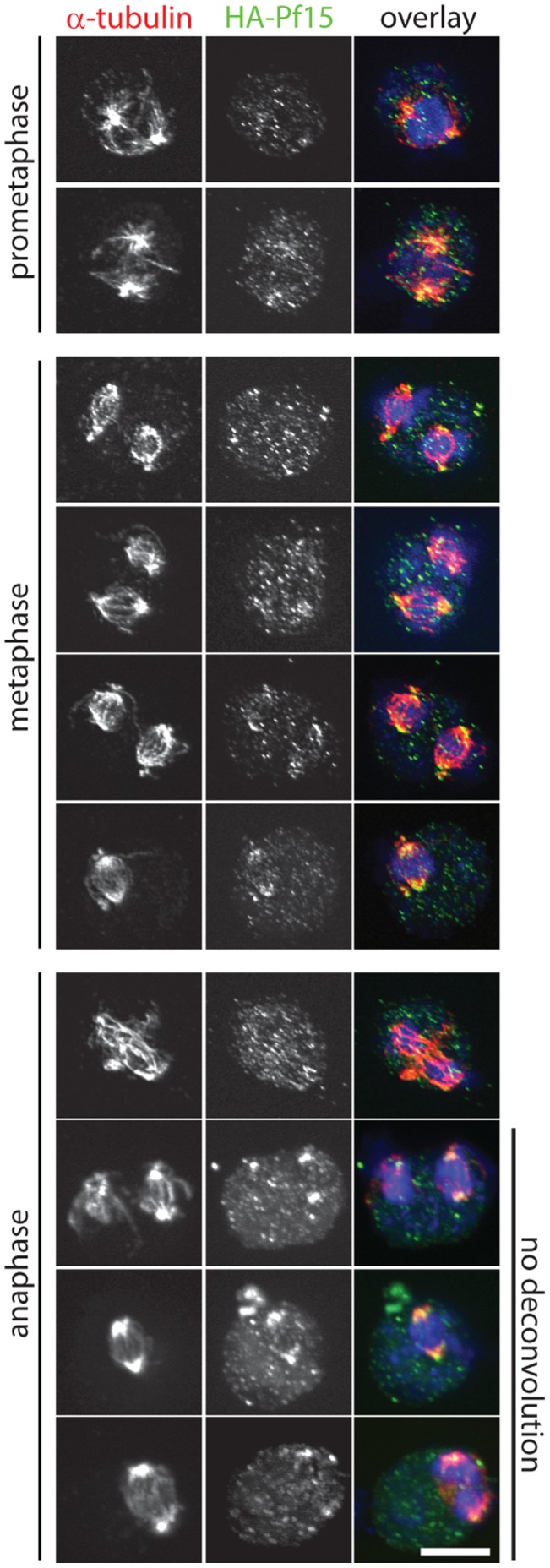
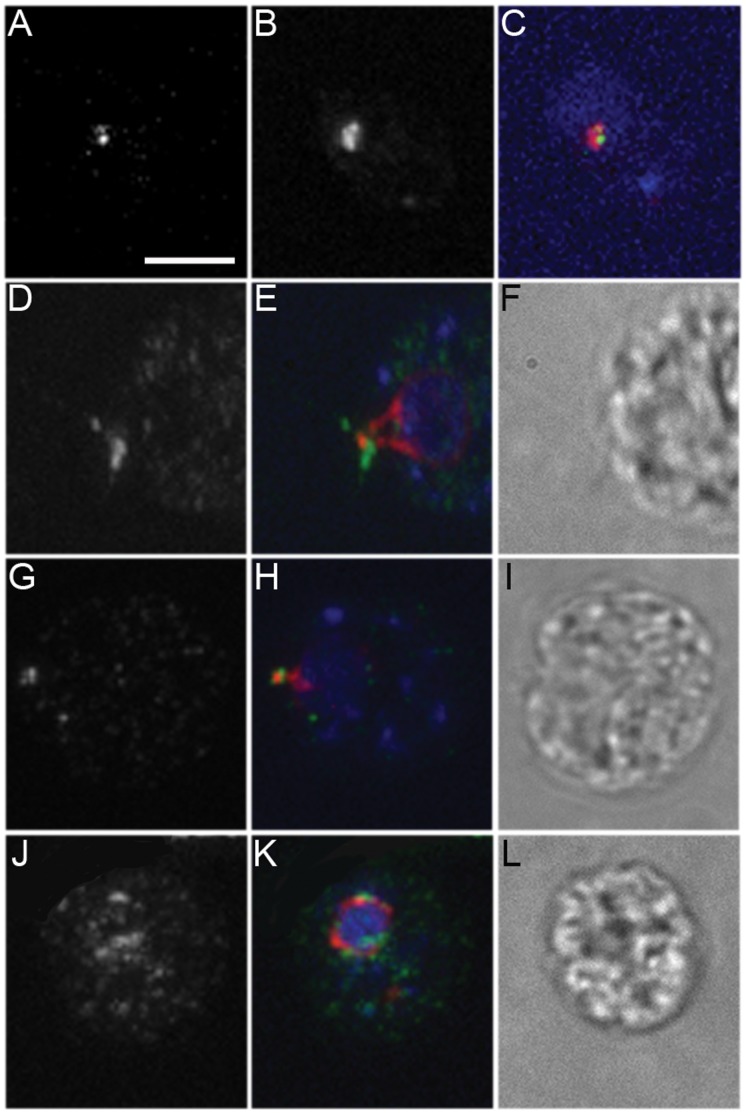
Centrioles and basal bodies are essential for a variety of cellular processes that include the recruitment of proteins to these structures for both centrosomal and ciliary function. This recruitment is compromised when centriole/basal body assembly is defective. Mutations that cause basal body assembly defects confer supersensitivity to Taxol. These include bld2, bld10, bld12, uni3, vfl1, vfl2, and vfl3. Flagellar motility mutants do not confer sensitivity with the exception of mutations in the p60 (pf19) and p80 (pf15) subunits of the microtubule severing protein katanin. We have identified additional pf15 and bld2 (ε-tubulin) alleles in screens for Taxol sensitivity. Null pf15 and bld2 alleles are viable and are not essential genes in Chlamydomonas. Analysis of double mutant strains with the pf15-3 and bld2-6 null alleles suggests that basal bodies in Chlamydomonas may recruit additional proteins beyond katanin that affect spindle microtubule stability. The bld2-5 allele is a hypomorphic allele and its phenotype is modulated by nutritional cues. Basal bodies in bld2-5 cells are missing proximal ends. The basal body mutants show aberrant localization of an epitope-tagged p80 subunit of katanin. Unlike IFT proteins, katanin p80 does not localize to the transition fibers of the basal bodies based on an analysis of the uni1 mutant as well as the lack of colocalization of katanin p80 with IFT74. We suggest that the triplet microtubules are likely to play a key role in katanin p80 recruitment to the basal body of Chlamydomonas rather than the transition fibers that are needed for IFT localization.
Conflict of interest statement
Figures
References
-
- Hartman JJ, Mahr J, McNally K, Okawa K, Iwamatsu A, et al. (1998) Katanin, a microtubule-severing protein, is a novel AAA ATPase that targets to the centrosome using a WD40-containing subunit. Cell 93: 277–287. - PubMed
-
- McNally FJ, Okawa K, Iwamatsu A, Vale RD (1996) Katanin, the microtubule-severing ATPase, is concentrated at centrosomes. J Cell Sci 109 (Pt 3): 561–567. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
