

The regulation of runt-related transcription factor 2 by fibroblast growth factor-2 and connexin43 requires the inositol polyphosphate/protein kinase Cδ cascade
- PMID: 23322705
- PMCID: PMC3657330
- DOI: 10.1002/jbmr.1867
The regulation of runt-related transcription factor 2 by fibroblast growth factor-2 and connexin43 requires the inositol polyphosphate/protein kinase Cδ cascade
Abstract
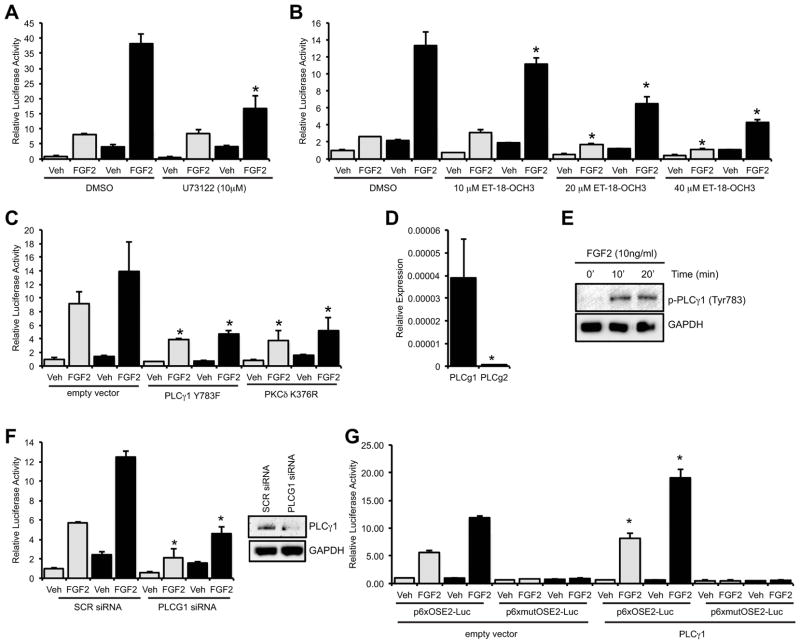
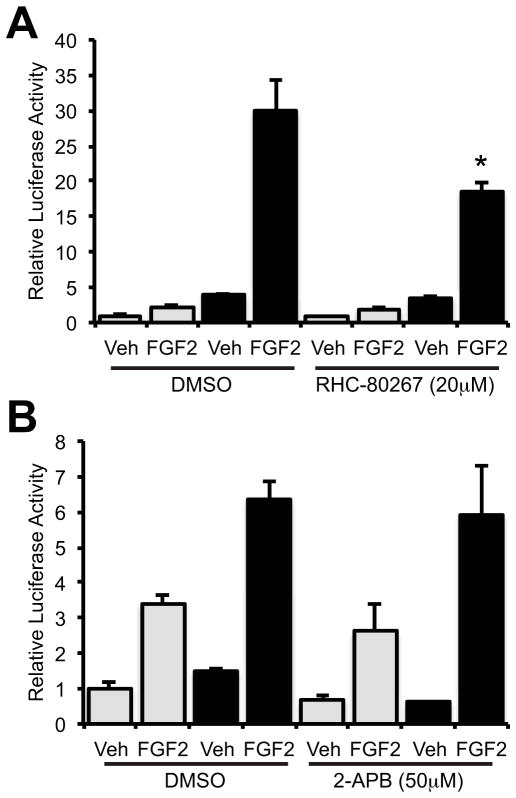
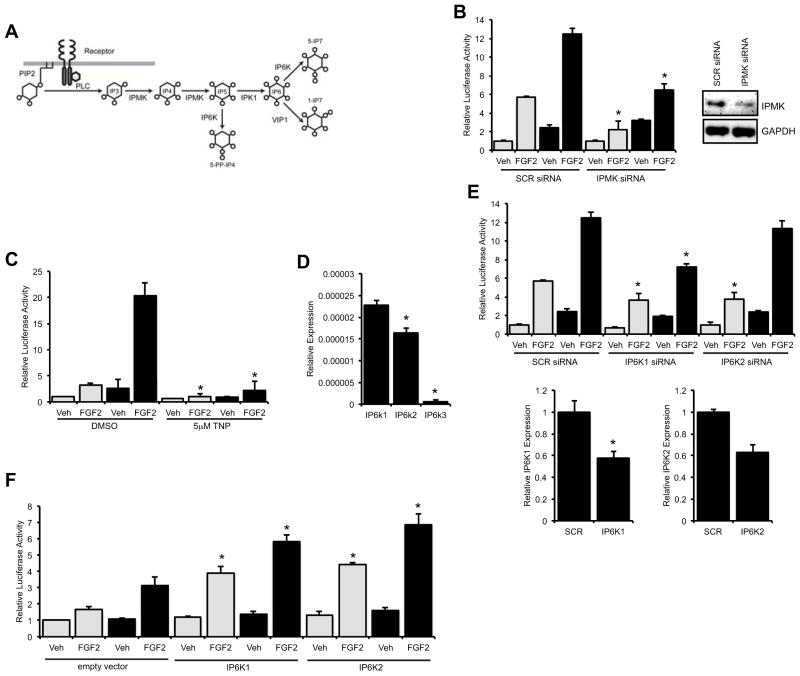
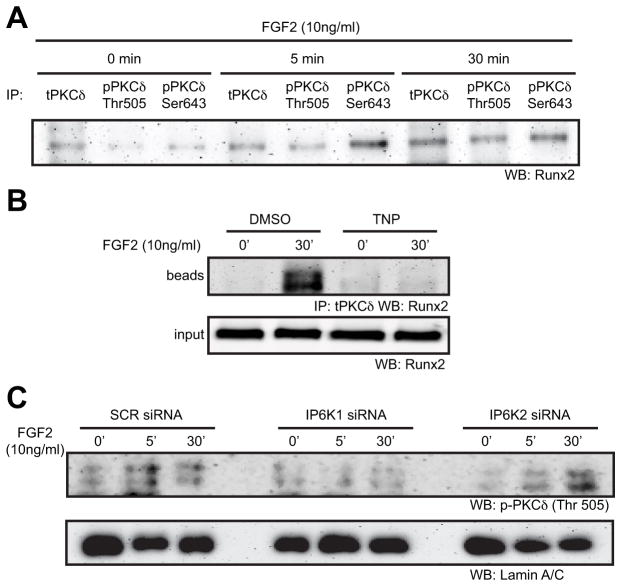
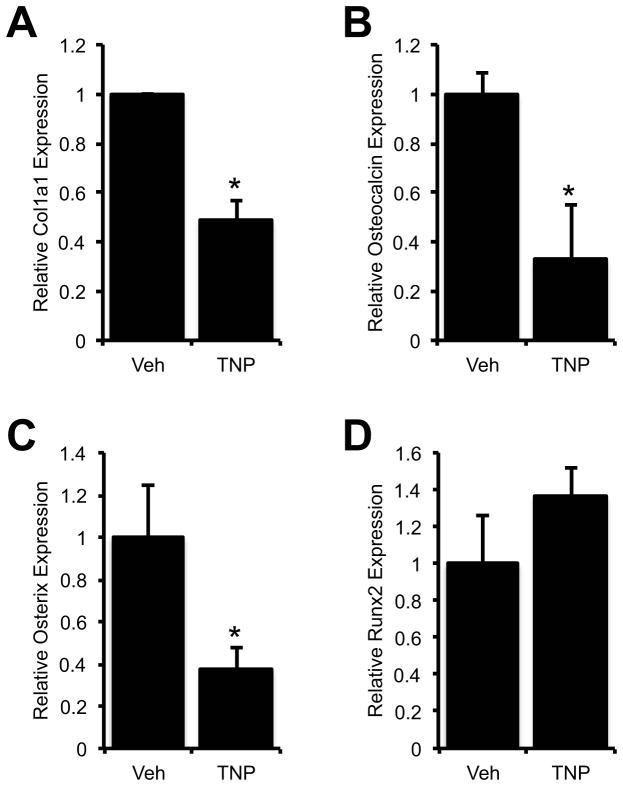
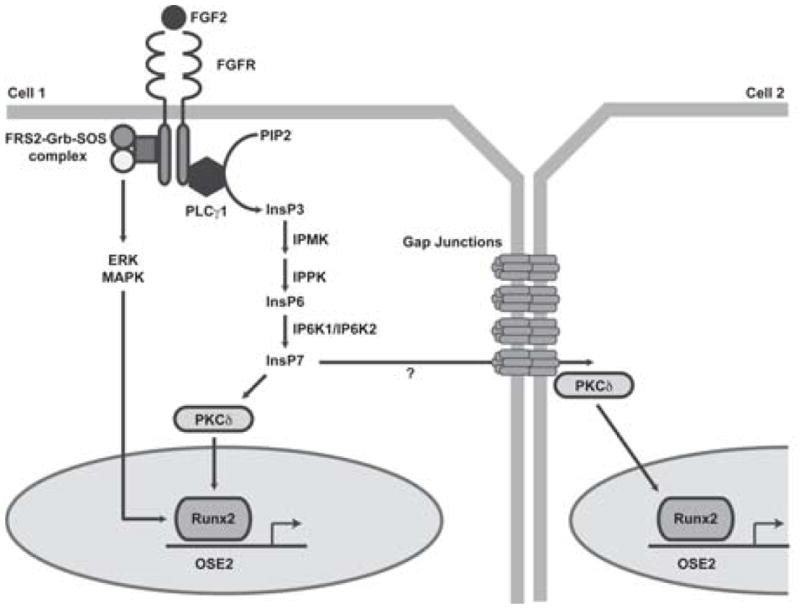
Connexin43 (Cx43) plays a critical role in osteoblast function and bone mass accrual, yet the identity of the second messengers communicated by Cx43 gap junctions, the targets of these second messengers and how they regulate osteoblast function remain largely unknown. We have shown that alterations of Cx43 expression in osteoblasts can impact the responsiveness to fibroblast growth factor-2 (FGF2), by modulating the transcriptional activity of runt-related transcription factor 2 (Runx2). In this study, we examined the contribution of the phospholipase Cγ1/inositol polyphosphate/protein kinase C delta (PKCδ) cascade to the Cx43-dependent transcriptional response of MC3T3 osteoblasts to FGF2. Knockdown of expression and/or inhibition of function of phospholipase Cγ1, inositol polyphosphate multikinase, which generates inositol 1,3,4,5-tetrakisphosphate (InsP₄) and InsP₅, and inositol hexakisphosphate kinase 1/2, which generates inositol pyrophosphates, prevented the ability of Cx43 to potentiate FGF2-induced signaling through Runx2. Conversely, overexpression of phospholipase Cγ1 and inositol hexakisphosphate kinase 1/2 enhanced FGF2 activation of Runx2 and the effect of Cx43 overexpression on this response. Disruption of these pathways blocked the nuclear accumulation of PKCδ and the FGF2-dependent interaction of PKCδ and Runx2, reducing Runx2 transcriptional activity. These data reveal that FGF2-signaling involves the inositol polyphosphate cascade, including inositol hexakisphosphate kinase (IP6K), and demonstrate that IP6K regulates Runx2 and osteoblast gene expression. Additionally, these data implicate the water-soluble inositol polyphosphates as mediators of the Cx43-dependent amplification of the osteoblast response to FGF2, and suggest that these low molecular weight second messengers may be biologically relevant mediators of osteoblast function that are communicated by Cx43-gap junctions.
Copyright © 2013 American Society for Bone and Mineral Research.
Conflict of interest statement
All authors state that they have no conflicts of interest.
Figures
Similar articles
-
ERK acts in parallel to PKCδ to mediate the connexin43-dependent potentiation of Runx2 activity by FGF2 in MC3T3 osteoblasts.Am J Physiol Cell Physiol. 2012 Apr 1;302(7):C1035-44. doi: 10.1152/ajpcell.00262.2011. Epub 2012 Jan 25. Am J Physiol Cell Physiol. 2012. PMID: 22277757 Free PMC article.
-
Connexin43 potentiates osteoblast responsiveness to fibroblast growth factor 2 via a protein kinase C-delta/Runx2-dependent mechanism.Mol Biol Cell. 2009 Jun;20(11):2697-708. doi: 10.1091/mbc.e08-10-1079. Epub 2009 Apr 1. Mol Biol Cell. 2009. PMID: 19339281 Free PMC article.
-
An intact connexin43 is required to enhance signaling and gene expression in osteoblast-like cells.J Cell Biochem. 2013 Nov;114(11):2542-50. doi: 10.1002/jcb.24603. J Cell Biochem. 2013. PMID: 23744706 Free PMC article.
-
Molecular mechanisms of osteoblast/osteocyte regulation by connexin43.Calcif Tissue Int. 2014 Jan;94(1):55-67. doi: 10.1007/s00223-013-9742-6. Epub 2013 Jun 11. Calcif Tissue Int. 2014. PMID: 23754488 Free PMC article. Review.
-
Inositol phosphate kinases: Expanding the biological significance of the universal core of the protein kinase fold.Adv Biol Regul. 2019 Jan;71:118-127. doi: 10.1016/j.jbior.2018.10.006. Epub 2018 Oct 27. Adv Biol Regul. 2019. PMID: 30392847 Free PMC article. Review.
Cited by
-
A cost-effective method to enhance adenoviral transduction of primary murine osteoblasts and bone marrow stromal cells.Bone Res. 2016 Aug 9;4:16021. doi: 10.1038/boneres.2016.21. eCollection 2016. Bone Res. 2016. PMID: 27547486 Free PMC article.
-
Deficiency of the intermediate filament synemin reduces bone mass in vivo.Am J Physiol Cell Physiol. 2016 Dec 1;311(6):C839-C845. doi: 10.1152/ajpcell.00218.2016. Epub 2016 Sep 7. Am J Physiol Cell Physiol. 2016. PMID: 27605453 Free PMC article.
-
Identification of shoulder osteoarthritis biomarkers: comparison between shoulders with and without osteoarthritis.J Shoulder Elbow Surg. 2015 Mar;24(3):382-90. doi: 10.1016/j.jse.2014.11.039. Epub 2015 Jan 13. J Shoulder Elbow Surg. 2015. PMID: 25595362 Free PMC article.
-
Connexins in the skeleton.Semin Cell Dev Biol. 2016 Feb;50:31-9. doi: 10.1016/j.semcdb.2015.12.017. Epub 2015 Dec 29. Semin Cell Dev Biol. 2016. PMID: 26740471 Free PMC article. Review.
-
Connexin43 Mediated Delivery of ADAMTS5 Targeting siRNAs from Mesenchymal Stem Cells to Synovial Fibroblasts.PLoS One. 2015 Jun 15;10(6):e0129999. doi: 10.1371/journal.pone.0129999. eCollection 2015. PLoS One. 2015. PMID: 26076025 Free PMC article.
References
-
- Chung DJ, Castro CH, Watkins M, Stains JP, Chung MY, Szejnfeld VL, Willecke K, Theis M, Civitelli R. Low peak bone mass and attenuated anabolic response to parathyroid hormone in mice with an osteoblast-specific deletion of connexin43. J Cell Sci. 2006;119(Pt 20):4187–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
