

Critical roles of type III phosphatidylinositol phosphate kinase in murine embryonic visceral endoderm and adult intestine
- PMID: 23322734
- PMCID: PMC3562790
- DOI: 10.1073/pnas.1213212110
Critical roles of type III phosphatidylinositol phosphate kinase in murine embryonic visceral endoderm and adult intestine
Abstract
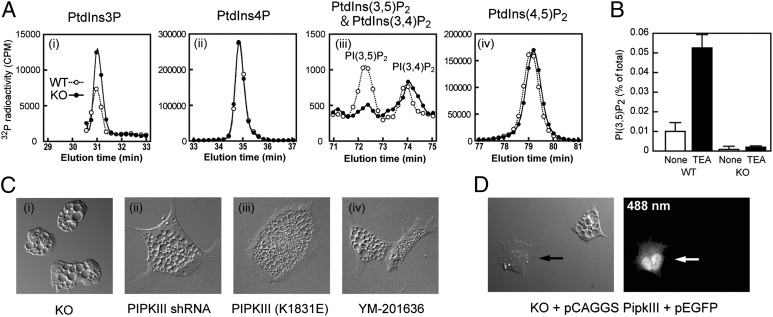
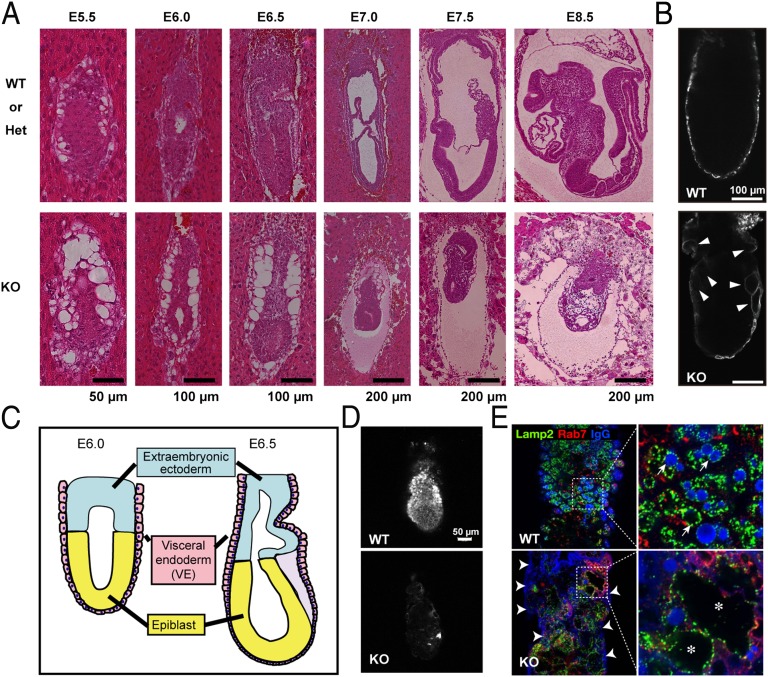
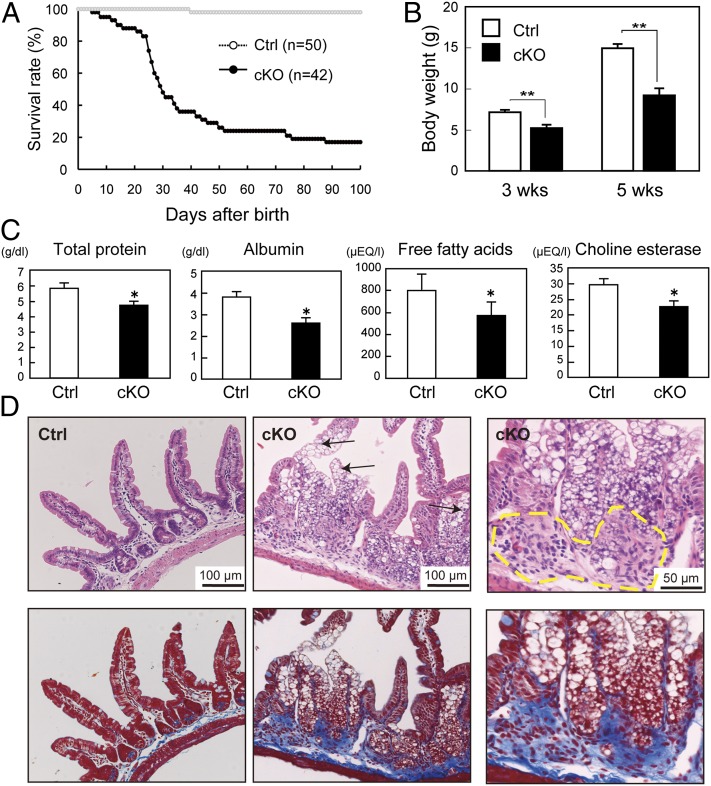
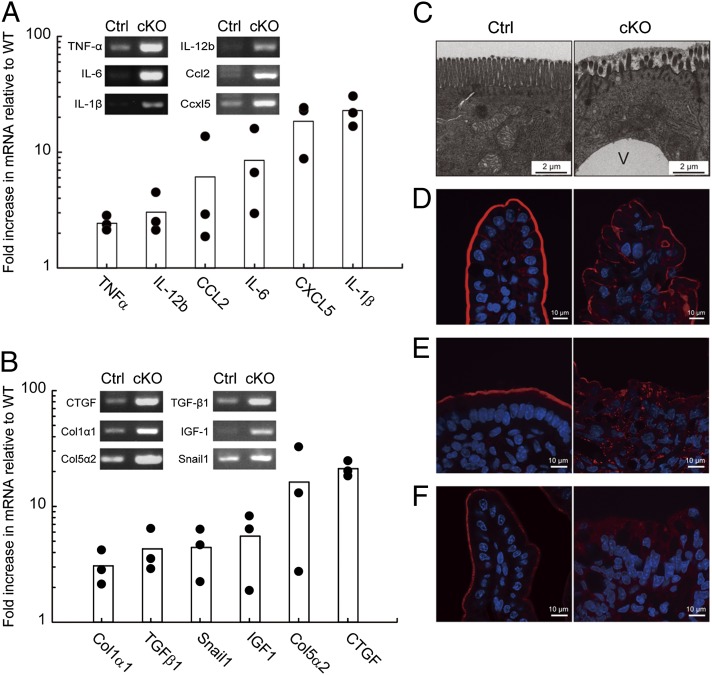
The metabolism of membrane phosphoinositides is critical for a variety of cellular processes. Phosphatidylinositol-3,5-bisphosphate [PtdIns(3,5)P(2)] controls multiple steps of the intracellular membrane trafficking system in both yeast and mammalian cells. However, other than in neuronal tissues, little is known about the physiological functions of PtdIns(3,5)P(2) in mammals. Here, we provide genetic evidence that type III phosphatidylinositol phosphate kinase (PIPKIII), which produces PtdIns(3,5)P(2), is essential for the functions of polarized epithelial cells. PIPKIII-null mouse embryos die by embryonic day 8.5 because of a failure of the visceral endoderm to supply the epiblast with maternal nutrients. Similarly, although intestine-specific PIPKIII-deficient mice are born, they fail to thrive and eventually die of malnutrition. At the mechanistic level, we show that PIPKIII regulates the trafficking of proteins to a cell's apical membrane domain. Importantly, mice with intestine-specific deletion of PIPKIII exhibit diarrhea and bloody stool, and their gut epithelial layers show inflammation and fibrosis, making our mutants an improved model for inflammatory bowel diseases. In summary, our data demonstrate that PIPKIII is required for the structural and functional integrity of two different types of polarized epithelial cells and suggest that PtdIns(3,5)P(2) metabolism is an unexpected and critical link between membrane trafficking in intestinal epithelial cells and the pathogenesis of inflammatory bowel disease.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Phosphatidylinositol-3,5-bisphosphate: metabolism and physiological functions.J Biochem. 2013 Sep;154(3):211-8. doi: 10.1093/jb/mvt064. Epub 2013 Jul 15. J Biochem. 2013. PMID: 23857703 Review.
-
Smg1 is required for embryogenesis and regulates diverse genes via alternative splicing coupled to nonsense-mediated mRNA decay.Proc Natl Acad Sci U S A. 2010 Jul 6;107(27):12186-91. doi: 10.1073/pnas.1007336107. Epub 2010 Jun 21. Proc Natl Acad Sci U S A. 2010. PMID: 20566848 Free PMC article.
-
Autotaxin/lysophospholipase D-mediated lysophosphatidic acid signaling is required to form distinctive large lysosomes in the visceral endoderm cells of the mouse yolk sac.J Biol Chem. 2009 Nov 27;284(48):33561-70. doi: 10.1074/jbc.M109.012716. Epub 2009 Oct 5. J Biol Chem. 2009. PMID: 19808661 Free PMC article.
-
Rac1-dependent collective cell migration is required for specification of the anterior-posterior body axis of the mouse.PLoS Biol. 2010 Aug 3;8(8):e1000442. doi: 10.1371/journal.pbio.1000442. PLoS Biol. 2010. PMID: 20689803 Free PMC article.
-
FYVE finger proteins as effectors of phosphatidylinositol 3-phosphate.Chem Phys Lipids. 1999 Apr;98(1-2):87-94. doi: 10.1016/s0009-3084(99)00021-3. Chem Phys Lipids. 1999. PMID: 10358931 Review.
Cited by
-
Vacuoles in mammals: a subcellular structure indispensable for early embryogenesis.Bioarchitecture. 2013 Jan-Feb;3(1):13-9. doi: 10.4161/bioa.24126. Bioarchitecture. 2013. PMID: 23572040 Free PMC article. Review.
-
Class III PI 3-kinase is the main source of PtdIns3P substrate and membrane recruitment signal for PIKfyve constitutive function in podocyte endomembrane homeostasis.Biochim Biophys Acta. 2015 May;1853(5):1240-50. doi: 10.1016/j.bbamcr.2015.01.008. Epub 2015 Jan 22. Biochim Biophys Acta. 2015. PMID: 25619930 Free PMC article.
-
PIP kinases: A versatile family that demands further therapeutic attention.Adv Biol Regul. 2023 Jan;87:100939. doi: 10.1016/j.jbior.2022.100939. Epub 2022 Dec 5. Adv Biol Regul. 2023. PMID: 36517396 Free PMC article.
-
PIKfyve Deficiency in Myeloid Cells Impairs Lysosomal Homeostasis in Macrophages and Promotes Systemic Inflammation in Mice.Mol Cell Biol. 2019 Oct 11;39(21):e00158-19. doi: 10.1128/MCB.00158-19. Print 2019 Nov 1. Mol Cell Biol. 2019. PMID: 31427458 Free PMC article.
-
A novel phosphoinositide kinase Fab1 regulates biosynthesis of pathogenic aflatoxin in Aspergillus flavus.Virulence. 2021 Dec;12(1):96-113. doi: 10.1080/21505594.2020.1859820. Virulence. 2021. PMID: 33315533 Free PMC article.
References
-
- Sasaki T, et al. Mammalian phosphoinositide kinases and phosphatases. Prog Lipid Res. 2009;48(6):307–343. - PubMed
-
- Wymann MP, Schneiter R. Lipid signalling in disease. Nat Rev Mol Cell Biol. 2008;9(2):162–176. - PubMed
-
- Di Paolo G, De Camilli P. Phosphoinositides in cell regulation and membrane dynamics. Nature. 2006;443(7112):651–657. - PubMed
-
- Dove SK, et al. Osmotic stress activates phosphatidylinositol-3,5-bisphosphate synthesis. Nature. 1997;390(6656):187–192. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
