

Status epilepticus enhances tonic GABA currents and depolarizes GABA reversal potential in dentate fast-spiking basket cells
- PMID: 23324316
- PMCID: PMC3628014
- DOI: 10.1152/jn.00891.2012
Status epilepticus enhances tonic GABA currents and depolarizes GABA reversal potential in dentate fast-spiking basket cells
Abstract
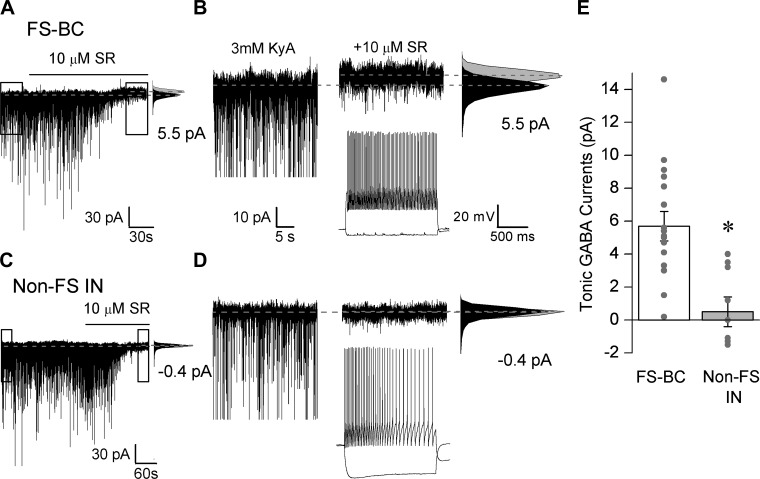
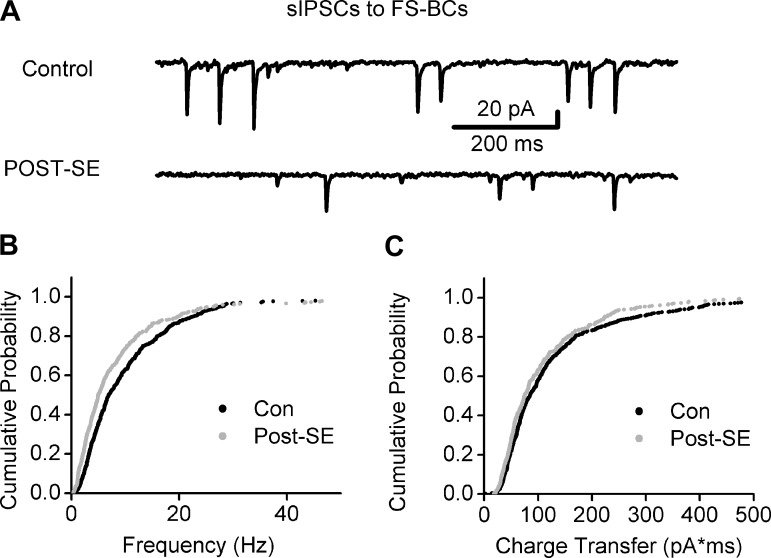
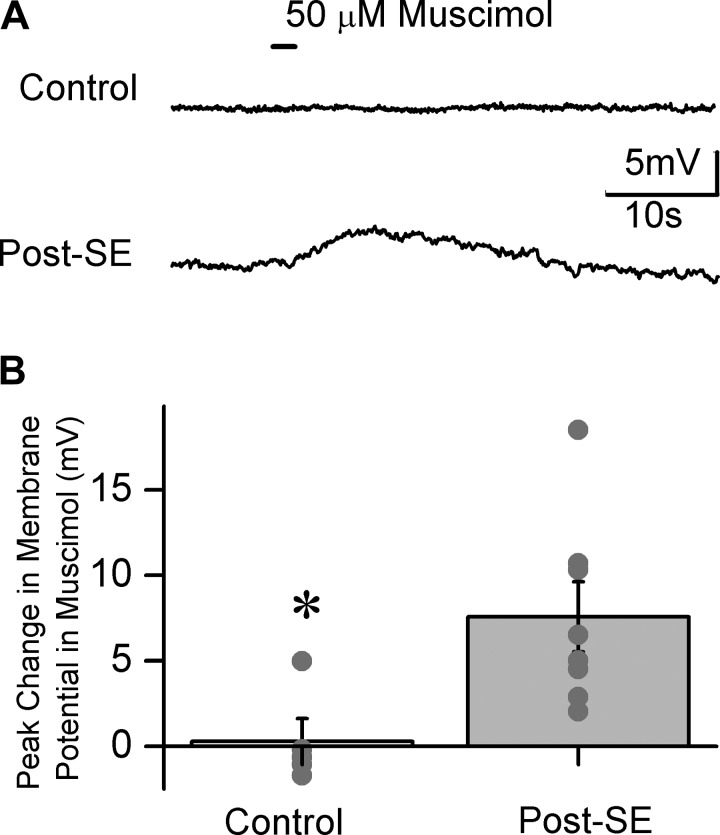
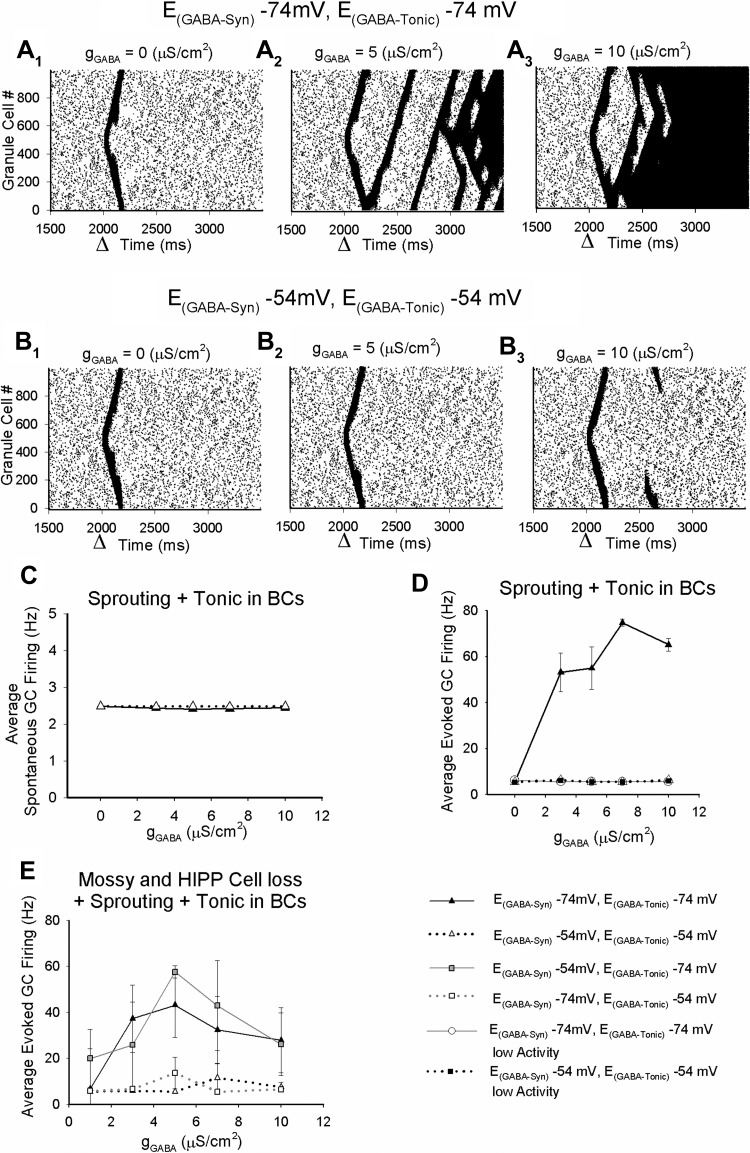
Temporal lobe epilepsy is associated with loss of interneurons and inhibitory dysfunction in the dentate gyrus. While status epilepticus (SE) leads to changes in granule cell inhibition, whether dentate basket cells critical for regulating granule cell feedforward and feedback inhibition express tonic GABA currents (I(GABA)) and undergo changes in inhibition after SE is not known. We find that interneurons immunoreactive for parvalbumin in the hilar-subgranular region express GABAA receptor (GABA(A)R) δ-subunits, which are known to underlie tonic I(GABA). Dentate fast-spiking basket cells (FS-BCs) demonstrate baseline tonic I(GABA) blocked by GABA(A)R antagonists. In morphologically and physiologically identified FS-BCs, tonic I(GABA) is enhanced 1 wk after pilocarpine-induced SE, despite simultaneous reduction in spontaneous inhibitory postsynaptic current (sIPSC) frequency. Amplitude of tonic I(GABA) in control and post-SE FS-BCs is enhanced by 4,5,6,7-tetrahydroisoxazolo[5,4-c]pyridin-3-ol (THIP), demonstrating the contribution of GABA(A)R δ-subunits. Whereas FS-BC resting membrane potential is unchanged after SE, perforated-patch recordings from FS-BCs show that the reversal potential for GABA currents (E(GABA)) is depolarized after SE. In model FS-BCs, increasing tonic GABA conductance decreased excitability when E(GABA) was shunting and increased excitability when E(GABA) was depolarizing. Although simulated focal afferent activation evoked seizurelike activity in model dentate networks with FS-BC tonic GABA conductance and shunting E(GABA), excitability of identical networks with depolarizing FS-BC E(GABA) showed lower activity levels. Thus, together, post-SE changes in tonic I(GABA) and E(GABA) maintain homeostasis of FS-BC activity and limit increases in dentate excitability. These findings have implications for normal FS-BC function and can inform studies examining comorbidities and therapeutics following SE.
Figures







References
-
- Arima-Yoshida F, Watabe AM, Manabe T. The mechanisms of the strong inhibitory modulation of long-term potentiation in the rat dentate gyrus. Eur J Neurosci 33: 1637–1646, 2011 - PubMed
-
- Baldi R, Varga C, Tamas G. Differential distribution of KCC2 along the axo-somato-dendritic axis of hippocampal principal cells. Eur J Neurosci 32: 1319–1325, 2010 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous