

Living without creatine: unchanged exercise capacity and response to chronic myocardial infarction in creatine-deficient mice
- PMID: 23325497
- PMCID: PMC4182017
- DOI: 10.1161/CIRCRESAHA.112.300725
Living without creatine: unchanged exercise capacity and response to chronic myocardial infarction in creatine-deficient mice
Abstract
Rationale: Creatine is thought to be involved in the spatial and temporal buffering of ATP in energetic organs such as heart and skeletal muscle. Creatine depletion affects force generation during maximal stimulation, while reduced levels of myocardial creatine are a hallmark of the failing heart, leading to the widely held view that creatine is important at high workloads and under conditions of pathological stress.
Objective: We therefore hypothesised that the consequences of creatine-deficiency in mice would be impaired running capacity, and exacerbation of heart failure following myocardial infarction.
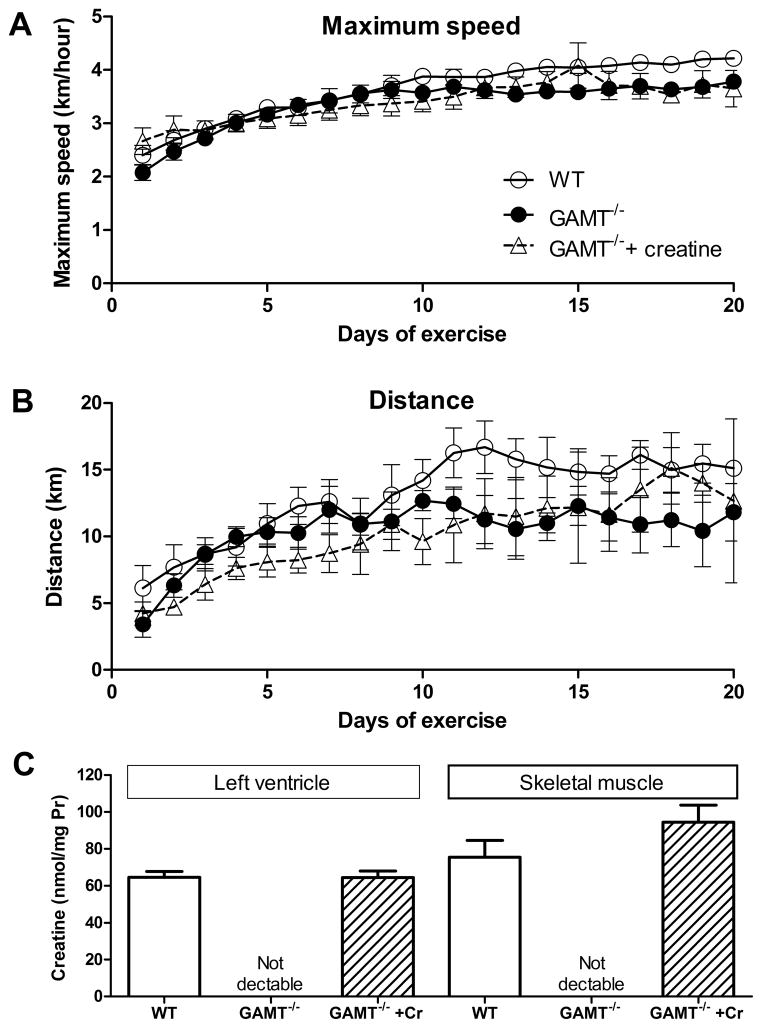
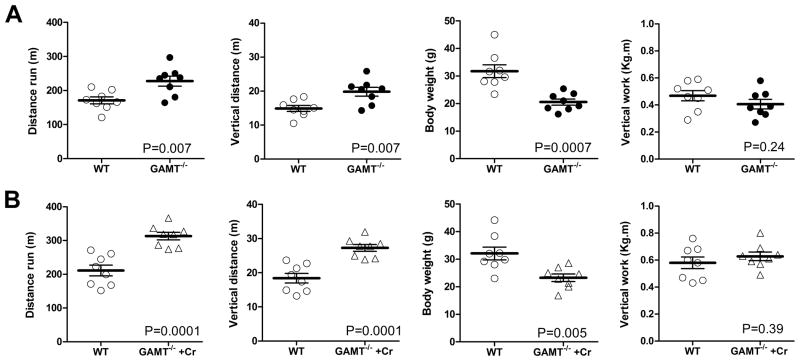
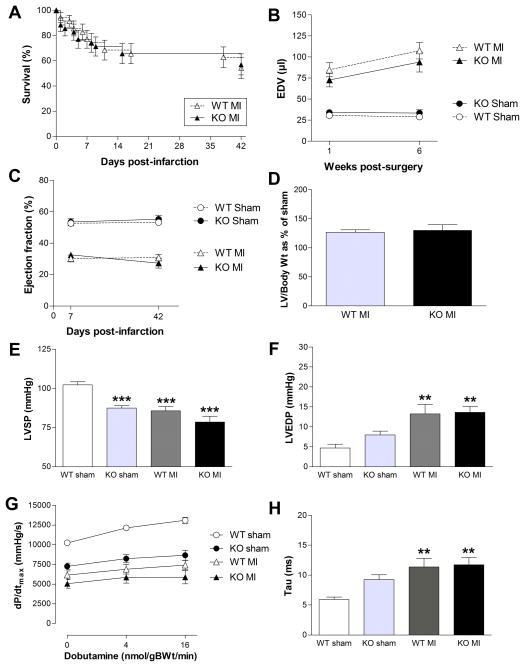
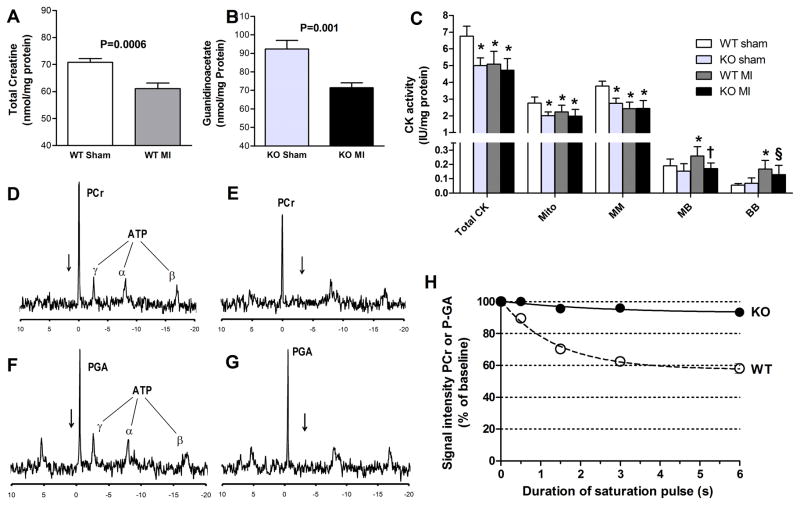
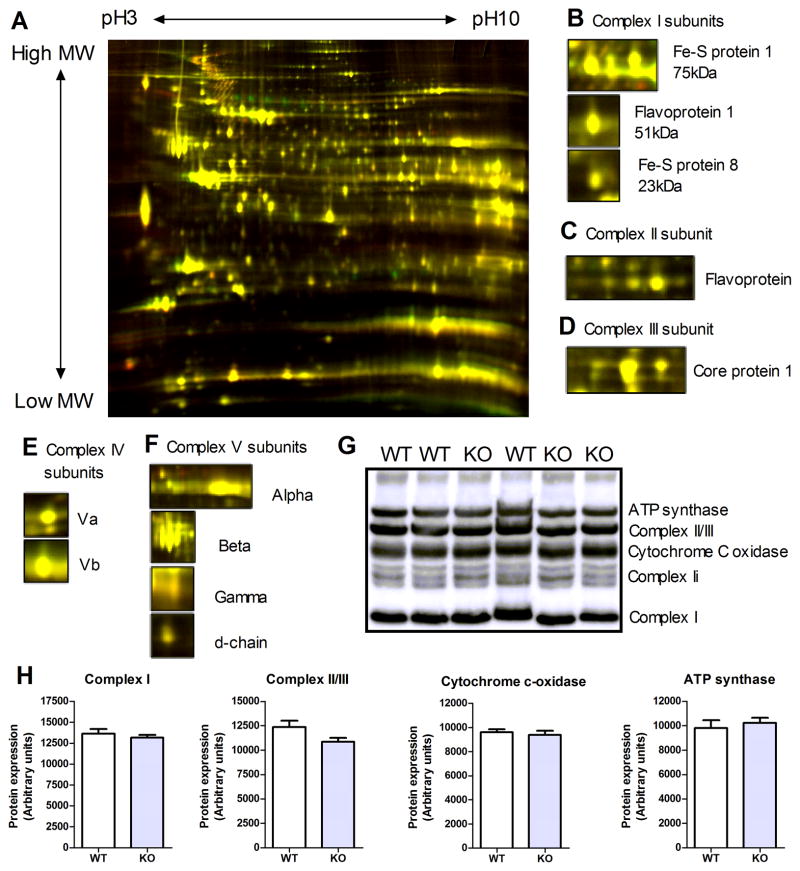
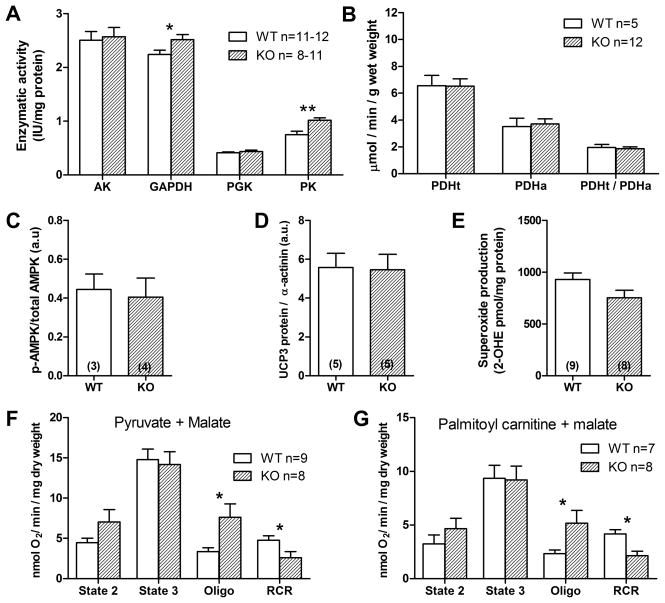
Methods and results: Surprisingly, mice with whole-body creatine deficiency due to knockout of the biosynthetic enzyme (guanidinoacetate N-methyltransferase [GAMT]) voluntarily ran just as fast and as far as controls (>10 km/night) and performed the same level of work when tested to exhaustion on a treadmill. Furthermore, survival following myocardial infarction was not altered, nor was subsequent left ventricular (LV) remodelling and development of chronic heart failure exacerbated, as measured by 3D-echocardiography and invasive hemodynamics. These findings could not be accounted for by compensatory adaptations, with no differences detected between WT and GAMT(-/-) proteomes. Alternative phosphotransfer mechanisms were explored; adenylate kinase activity was unaltered, and although GAMT(-/-) hearts accumulated the creatine precursor guanidinoacetate, this had negligible energy-transfer activity, while mitochondria retained near normal function.
Conclusions: Creatine-deficient mice show unaltered maximal exercise capacity and response to chronic myocardial infarction, and no obvious metabolic adaptations. Our results question the paradigm that creatine is essential for high workload and chronic stress responses in heart and skeletal muscle.
Figures
Comment in
-
Creatine--a dispensable metabolite?Circ Res. 2013 Mar 15;112(6):878-80. doi: 10.1161/CIRCRESAHA.113.300974. Circ Res. 2013. PMID: 23493302 Free PMC article. No abstract available.
Similar articles
-
Cardiac expression and location of hexokinase changes in a mouse model of pure creatine deficiency.Am J Physiol Heart Circ Physiol. 2021 Feb 1;320(2):H613-H629. doi: 10.1152/ajpheart.00188.2020. Epub 2020 Dec 18. Am J Physiol Heart Circ Physiol. 2021. PMID: 33337958
-
Reduced inotropic reserve and increased susceptibility to cardiac ischemia/reperfusion injury in phosphocreatine-deficient guanidinoacetate-N-methyltransferase-knockout mice.Circulation. 2005 May 17;111(19):2477-85. doi: 10.1161/01.CIR.0000165147.99592.01. Epub 2005 May 9. Circulation. 2005. PMID: 15883212
-
Unchanged mitochondrial organization and compartmentation of high-energy phosphates in creatine-deficient GAMT-/- mouse hearts.Am J Physiol Heart Circ Physiol. 2013 Aug 15;305(4):H506-20. doi: 10.1152/ajpheart.00919.2012. Epub 2013 Jun 21. Am J Physiol Heart Circ Physiol. 2013. PMID: 23792673 Free PMC article.
-
AGAT, GAMT and SLC6A8 distribution in the central nervous system, in relation to creatine deficiency syndromes: a review.J Inherit Metab Dis. 2008 Apr;31(2):230-9. doi: 10.1007/s10545-008-0826-9. Epub 2008 Apr 4. J Inherit Metab Dis. 2008. PMID: 18392746 Review.
-
Clinical aspects of left ventricular diastolic function assessed by Doppler echocardiography following acute myocardial infarction.Dan Med Bull. 2001 Nov;48(4):199-210. Dan Med Bull. 2001. PMID: 11767125 Review.
Cited by
-
Creatine--a dispensable metabolite?Circ Res. 2013 Mar 15;112(6):878-80. doi: 10.1161/CIRCRESAHA.113.300974. Circ Res. 2013. PMID: 23493302 Free PMC article. No abstract available.
-
Myocardial creatine levels do not influence response to acute oxidative stress in isolated perfused heart.PLoS One. 2014 Oct 1;9(10):e109021. doi: 10.1371/journal.pone.0109021. eCollection 2014. PLoS One. 2014. PMID: 25272153 Free PMC article.
-
Role of Creatine in the Heart: Health and Disease.Nutrients. 2021 Apr 7;13(4):1215. doi: 10.3390/nu13041215. Nutrients. 2021. PMID: 33917009 Free PMC article. Review.
-
Mitochondrial Calcium Increase Induced by RyR1 and IP3R Channel Activation After Membrane Depolarization Regulates Skeletal Muscle Metabolism.Front Physiol. 2018 Jun 25;9:791. doi: 10.3389/fphys.2018.00791. eCollection 2018. Front Physiol. 2018. PMID: 29988564 Free PMC article.
-
Advanced physiological roles of guanidinoacetic acid.Eur J Nutr. 2015 Dec;54(8):1211-5. doi: 10.1007/s00394-015-1050-7. Epub 2015 Sep 28. Eur J Nutr. 2015. PMID: 26411433 Review.
References
-
- Brosnan J, da Silva R, Brosnan M. The metabolic burden of creatine synthesis. Amino Acids. 2011;40:1325–1331. - PubMed
-
- Ellington WR. Evolution and physiological roles of phosphagen systems. Annu Rev Physiol. 2001;63:289–325. - PubMed
-
- Ingwall JS, Weiss RG. Is the failing heart energy starved? On using chemical energy to support cardiac function. Circ Res. 2004;95:135–145. - PubMed
-
- Schlattner U, Tokarska-Schlattner M, Wallimann T. Mitochondrial creatine kinase in human health and disease. Biochim Biophys Acta. 2006;1762:164–180. - PubMed
-
- Brosnan JT, Brosnan ME. Creatine: endogenous metabolite, dietary, and therapeutic supplement. Annu Rev Nutr. 2007;27:241–261. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
