

Notch cooperates with Lozenge/Runx to lock haemocytes into a differentiation programme
- PMID: 23325760
- PMCID: PMC3557782
- DOI: 10.1242/dev.086785
Notch cooperates with Lozenge/Runx to lock haemocytes into a differentiation programme
Abstract
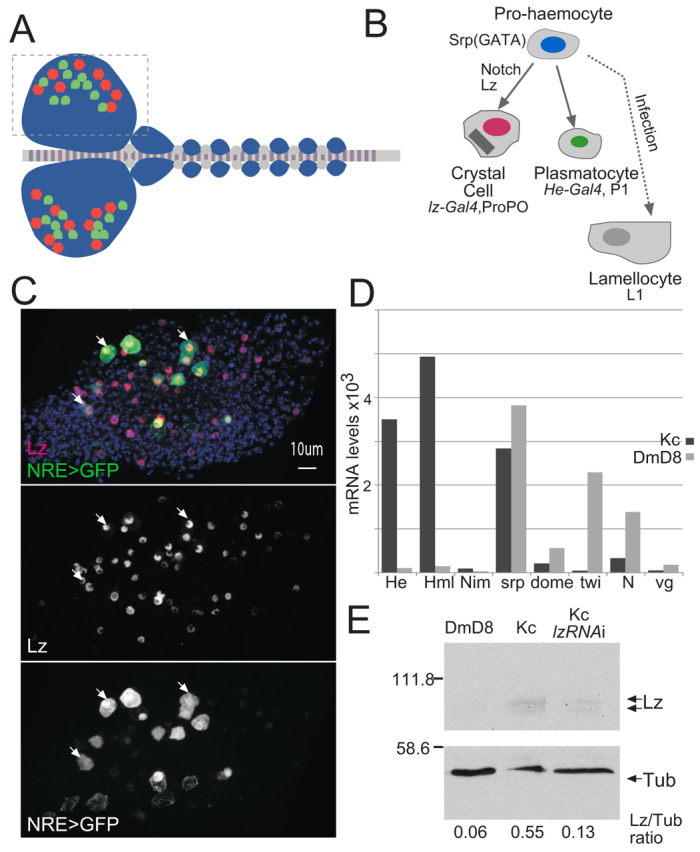
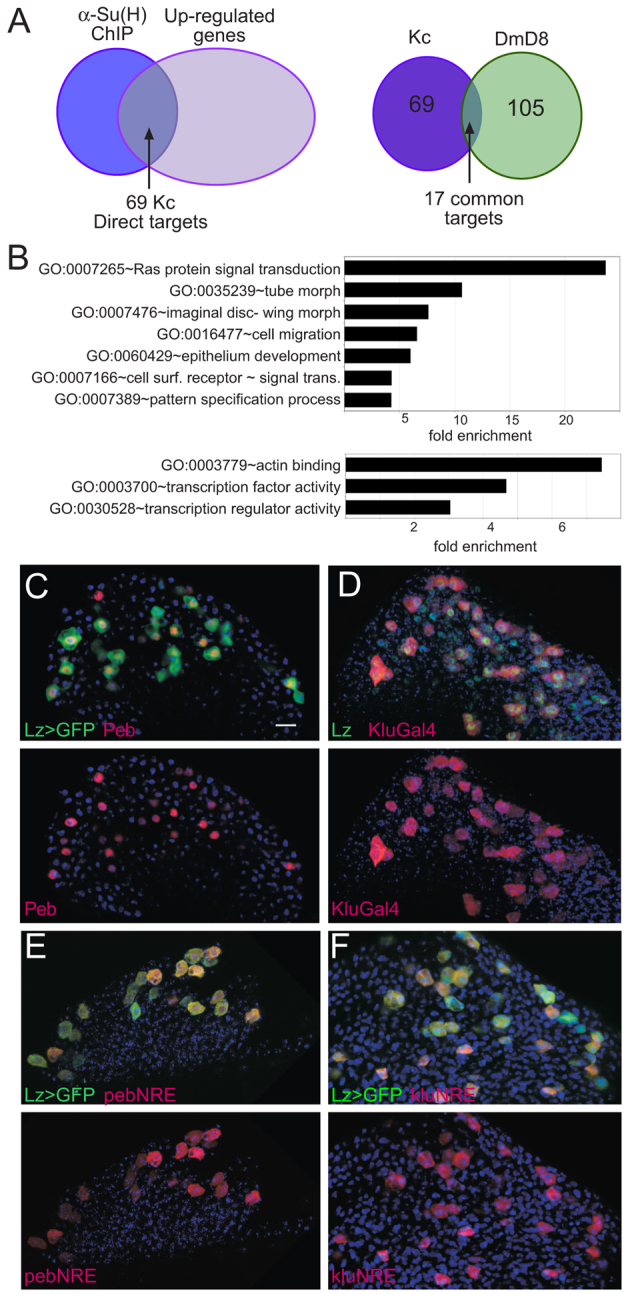
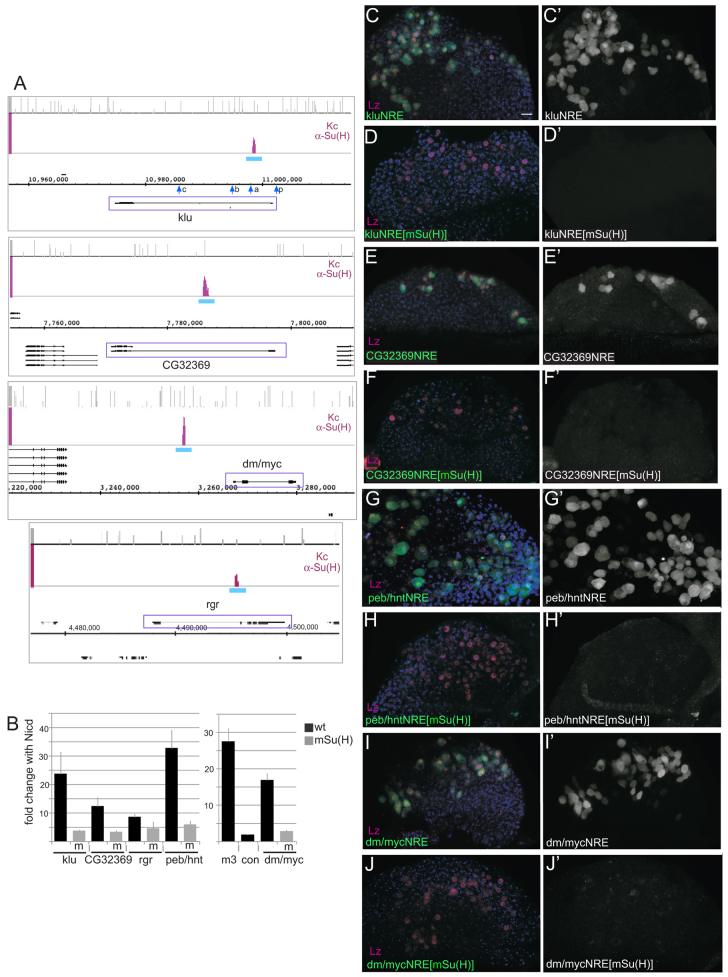
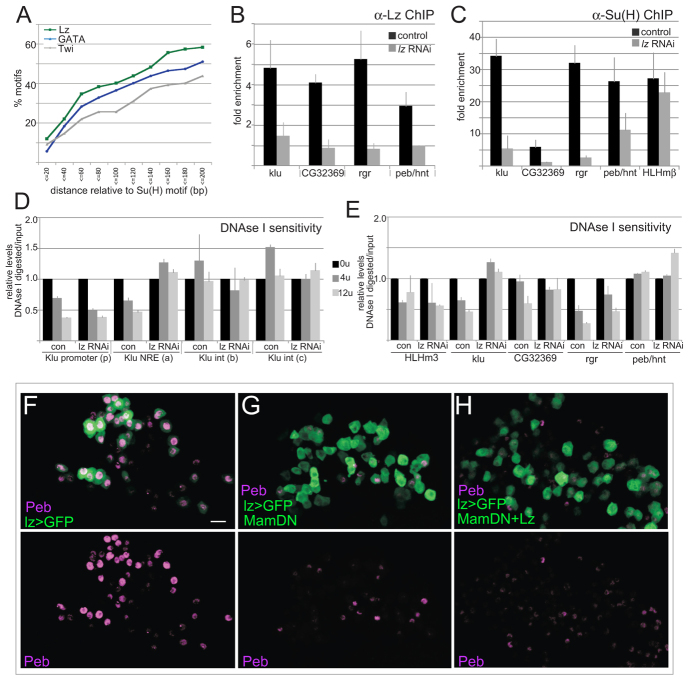
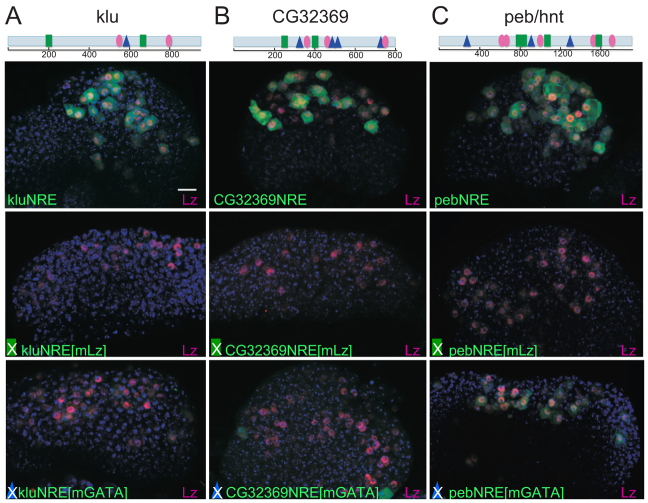
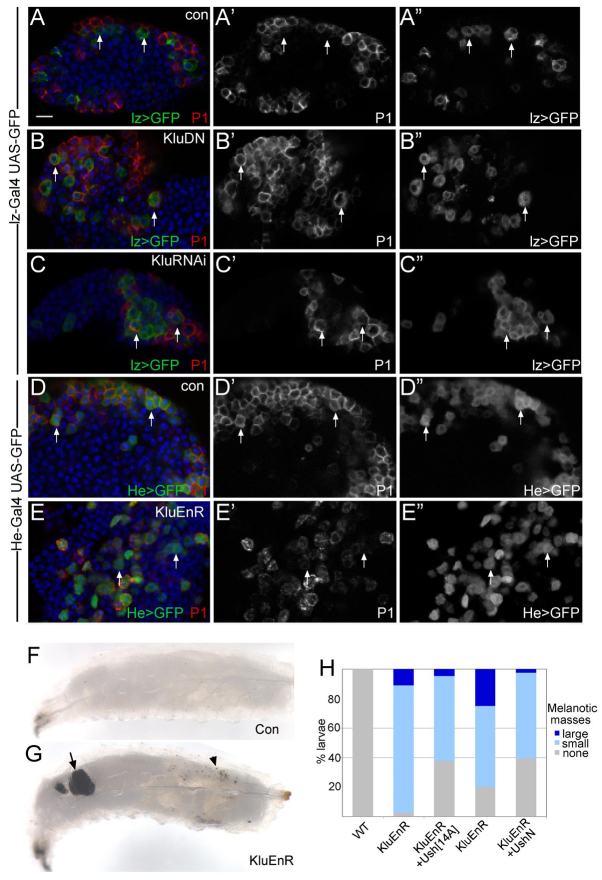
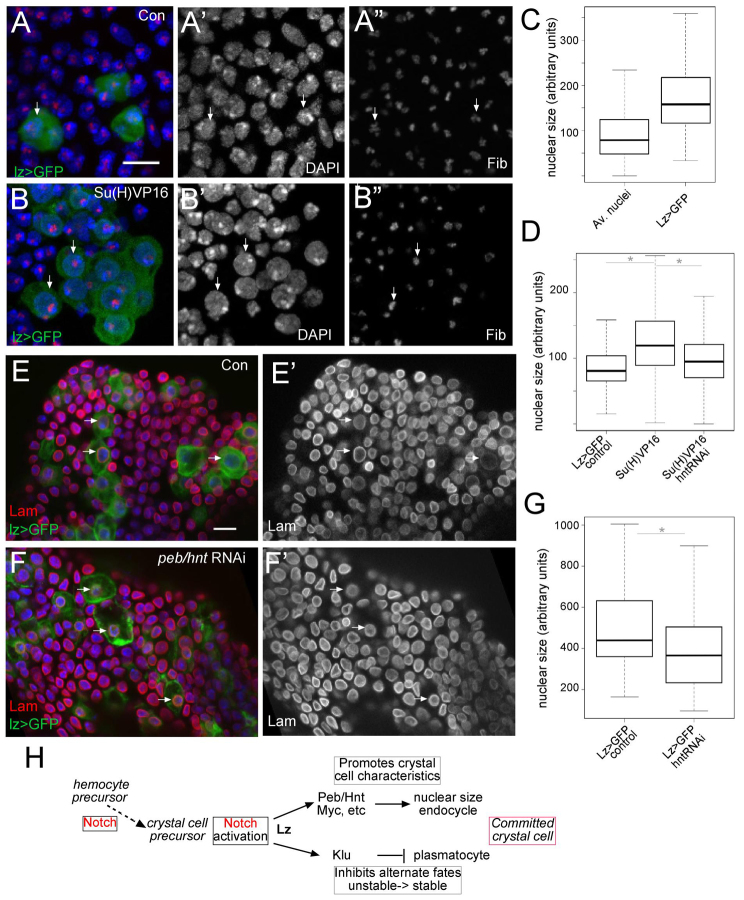
The diverse functions of Notch signalling imply that it must elicit context-specific programmes of gene expression. With the aim of investigating how Notch drives cells to differentiate, we have used a genome-wide approach to identify direct Notch targets in Drosophila haemocytes (blood cells), where Notch promotes crystal cell differentiation. Many of the identified Notch-regulated enhancers contain Runx and GATA motifs, and we demonstrate that binding of the Runx protein Lozenge (Lz) is required for enhancers to be competent to respond to Notch. Functional studies of targets, such as klumpfuss (ERG/WT1 family) and pebbled/hindsight (RREB1 homologue), show that Notch acts both to prevent the cells adopting alternate cell fates and to promote morphological characteristics associated with crystal cell differentiation. Inappropriate activity of Klumpfuss perturbs the differentiation programme, resulting in melanotic tumours. Thus, by acting as a master regulator, Lz directs Notch to activate selectively a combination of target genes that correctly locks cells into the differentiation programme.
Figures
Similar articles
-
A GATA/RUNX cis-regulatory module couples Drosophila blood cell commitment and differentiation into crystal cells.Dev Biol. 2007 May 15;305(2):726-34. doi: 10.1016/j.ydbio.2007.03.010. Epub 2007 Mar 13. Dev Biol. 2007. PMID: 17418114
-
A genome-wide RNA interference screen identifies a differential role of the mediator CDK8 module subunits for GATA/ RUNX-activated transcription in Drosophila.Mol Cell Biol. 2010 Jun;30(11):2837-48. doi: 10.1128/MCB.01625-09. Epub 2010 Apr 5. Mol Cell Biol. 2010. PMID: 20368357 Free PMC article.
-
Control of RUNX-induced repression of Notch signaling by MLF and its partner DnaJ-1 during Drosophila hematopoiesis.PLoS Genet. 2017 Jul 25;13(7):e1006932. doi: 10.1371/journal.pgen.1006932. eCollection 2017 Jul. PLoS Genet. 2017. PMID: 28742844 Free PMC article.
-
Notch signalling in the nucleus: roles of Mastermind-like (MAML) transcriptional coactivators.J Biochem. 2016 Mar;159(3):287-94. doi: 10.1093/jb/mvv123. Epub 2015 Dec 28. J Biochem. 2016. PMID: 26711237 Review.
-
Runx transcription factors and the developmental balance between cell proliferation and differentiation.Cell Biol Int. 2003;27(4):315-24. doi: 10.1016/s1065-6995(03)00018-0. Cell Biol Int. 2003. PMID: 12788047 Review.
Cited by
-
Drosophila as a Genetic Model for Hematopoiesis.Genetics. 2019 Feb;211(2):367-417. doi: 10.1534/genetics.118.300223. Genetics. 2019. PMID: 30733377 Free PMC article. Review.
-
De Novo DNA Synthesis in Aedes aegypti Midgut Cells as a Complementary Strategy to Limit Dengue Viral Replication.Front Microbiol. 2018 Apr 26;9:801. doi: 10.3389/fmicb.2018.00801. eCollection 2018. Front Microbiol. 2018. PMID: 29755433 Free PMC article.
-
The Role of Lozenge in Drosophila Hematopoiesis.Mol Cells. 2020 Feb 29;43(2):114-120. doi: 10.14348/molcells.2019.0249. Mol Cells. 2020. PMID: 31992020 Free PMC article. Review.
-
Identification of Cdca7 as a novel Notch transcriptional target involved in hematopoietic stem cell emergence.J Exp Med. 2014 Nov 17;211(12):2411-23. doi: 10.1084/jem.20131857. Epub 2014 Nov 10. J Exp Med. 2014. PMID: 25385755 Free PMC article.
-
Single-cell analysis of mosquito hemocytes identifies signatures of immune cell subtypes and cell differentiation.Elife. 2021 Jul 28;10:e66192. doi: 10.7554/eLife.66192. Elife. 2021. PMID: 34318744 Free PMC article.
References
-
- Alberta J. A., Springett G. M., Rayburn H., Natoli T. A., Loring J., Kreidberg J. A., Housman D. (2003). Role of the WT1 tumor suppressor in murine hematopoiesis. Blood 101, 2570-2574 - PubMed
-
- Benmimoun B., Polesello C., Waltzer L., Haenlin M. (2012). Dual role for Insulin/TOR signaling in the control of hematopoietic progenitor maintenance in Drosophila. Development 139, 1713-1717 - PubMed
-
- Bray S. J. (1997). Expression and function of Enhancer of split bHLH proteins during Drosophila neurogenesis. Perspect. Dev. Neurobiol. 4, 313-323 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
