

Nap1 regulates proper CENP-B binding to nucleosomes
- PMID: 23325853
- PMCID: PMC3597661
- DOI: 10.1093/nar/gks1464
Nap1 regulates proper CENP-B binding to nucleosomes
Abstract
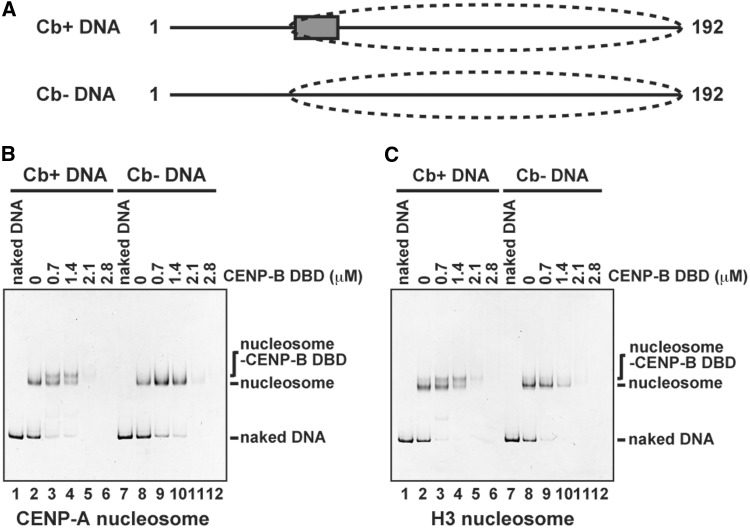
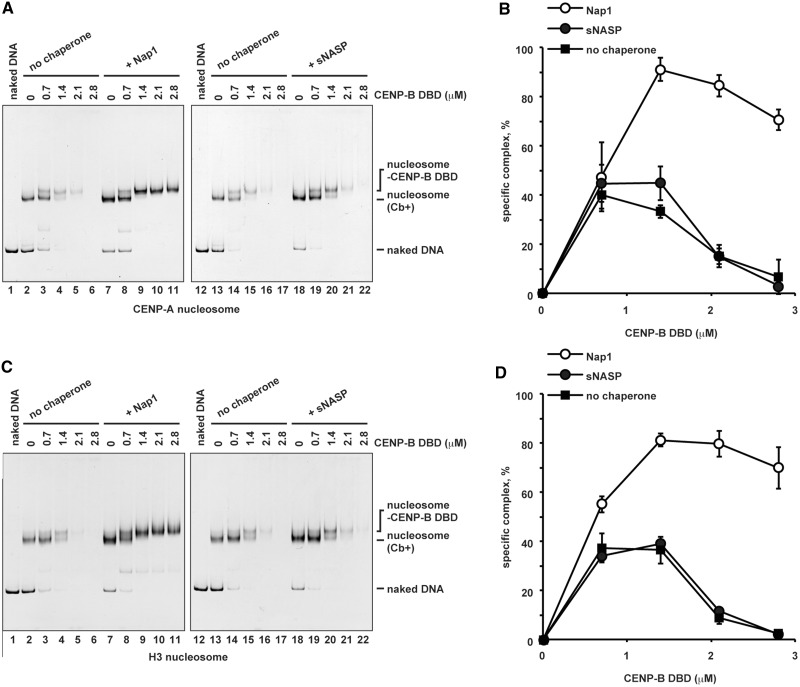
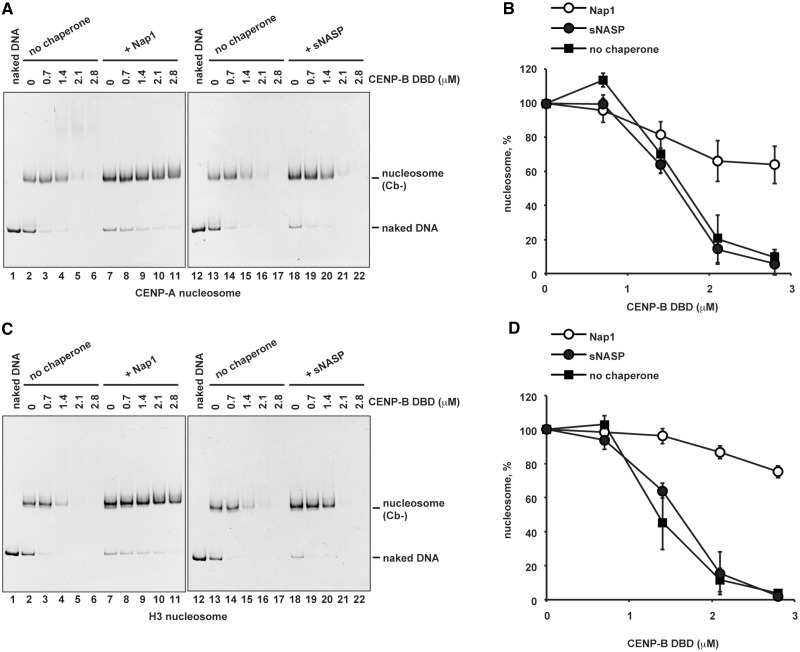
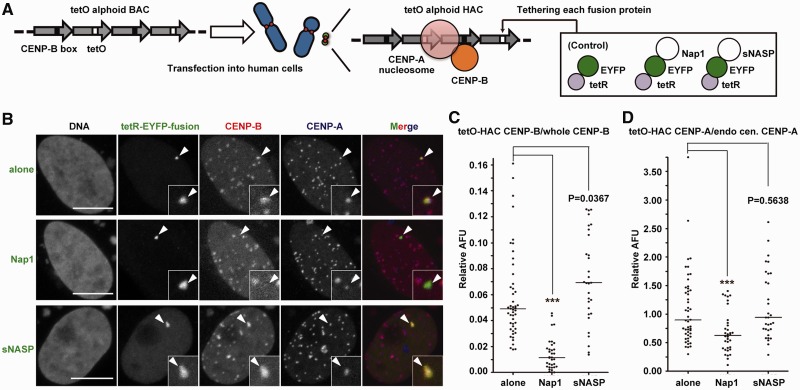
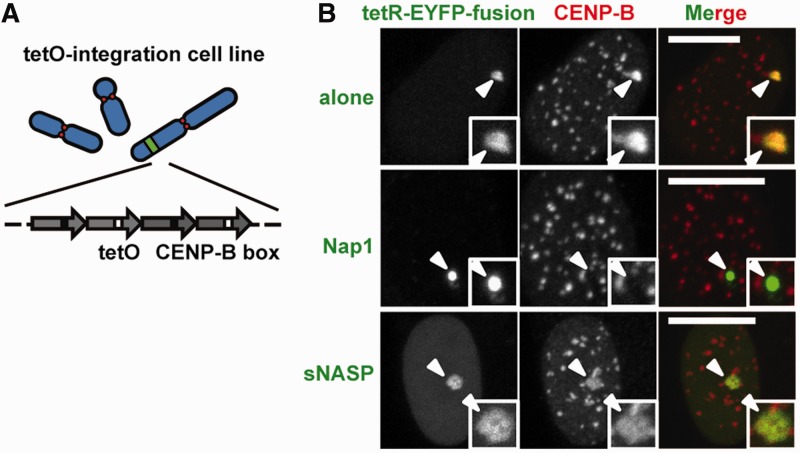
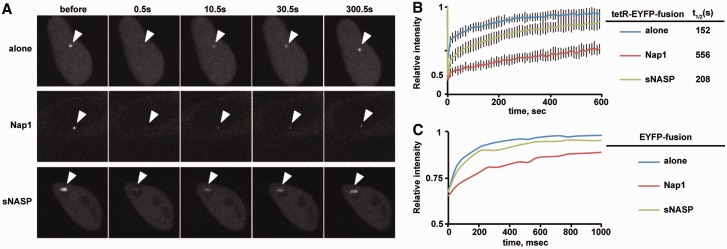
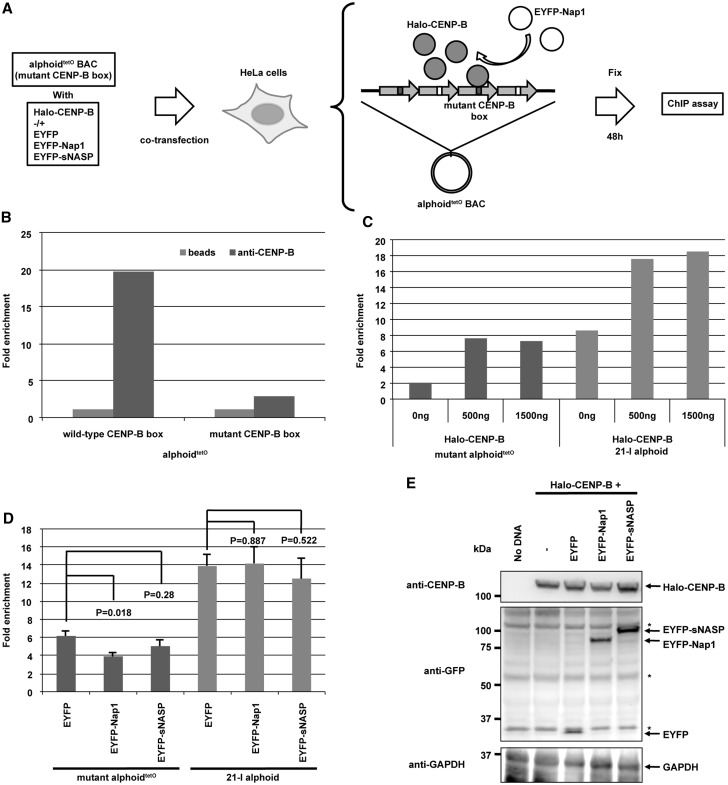
CENP-B is a widely conserved centromeric satellite DNA-binding protein, which specifically binds to a 17-bp DNA sequence known as the CENP-B box. CENP-B functions positively in the de novo assembly of centromeric nucleosomes, containing the centromere-specific histone H3 variant, CENP-A. At the same time, CENP-B also prevents undesired assembly of the CENP-A nucleosome through heterochromatin formation on satellite DNA integrated into ectopic sites. Therefore, improper CENP-B binding to chromosomes could be harmful. However, no CENP-B eviction mechanism has yet been reported. In the present study, we found that human Nap1, an acidic histone chaperone, inhibited the non-specific binding of CENP-B to nucleosomes and apparently stimulated CENP-B binding to its cognate CENP-B box DNA in nucleosomes. In human cells, the CENP-B eviction activity of Nap1 was confirmed in model experiments, in which the CENP-B binding to a human artificial chromosome or an ectopic chromosome locus bearing CENP-B boxes was significantly decreased when Nap1 was tethered near the CENP-B box sequence. In contrast, another acidic histone chaperone, sNASP, did not promote CENP-B eviction in vitro and in vivo and did not stimulate specific CENP-B binding to CENP-A nucleosomes in vitro. We therefore propose a novel mechanism of CENP-B regulation by Nap1.
Figures
Similar articles
-
Human centromere protein B induces translational positioning of nucleosomes on alpha-satellite sequences.J Biol Chem. 2005 Dec 16;280(50):41609-18. doi: 10.1074/jbc.M509666200. Epub 2005 Sep 23. J Biol Chem. 2005. PMID: 16183641
-
Comparison between the CENP-A and histone H3 structures in nucleosomes.Nucleus. 2012 Jan-Feb;3(1):6-11. doi: 10.4161/nucl.18372. Nucleus. 2012. PMID: 22127263
-
Stable complex formation of CENP-B with the CENP-A nucleosome.Nucleic Acids Res. 2015 May 26;43(10):4909-22. doi: 10.1093/nar/gkv405. Epub 2015 Apr 27. Nucleic Acids Res. 2015. PMID: 25916850 Free PMC article.
-
Putting CENP-A in its place.Cell Mol Life Sci. 2013 Feb;70(3):387-406. doi: 10.1007/s00018-012-1048-8. Epub 2012 Jun 23. Cell Mol Life Sci. 2013. PMID: 22729156 Free PMC article. Review.
-
Structure of the CENP-A nucleosome and its implications for centromeric chromatin architecture.Genes Genet Syst. 2011;86(6):357-64. doi: 10.1266/ggs.86.357. Genes Genet Syst. 2011. PMID: 22451475 Review.
Cited by
-
Diverse mechanisms of centromere specification.Curr Biol. 2021 Nov 22;31(22):R1491-R1504. doi: 10.1016/j.cub.2021.09.083. Curr Biol. 2021. PMID: 34813757 Free PMC article. Review.
-
Insight into G-quadruplex-hemin DNAzyme/RNAzyme: adjacent adenine as the intramolecular species for remarkable enhancement of enzymatic activity.Nucleic Acids Res. 2016 Sep 6;44(15):7373-84. doi: 10.1093/nar/gkw634. Epub 2016 Jul 15. Nucleic Acids Res. 2016. PMID: 27422869 Free PMC article.
-
Chromatin dynamics: H3K4 methylation and H3 variant replacement during development and in cancer.Cell Mol Life Sci. 2014 Sep;71(18):3439-63. doi: 10.1007/s00018-014-1605-4. Epub 2014 Mar 28. Cell Mol Life Sci. 2014. PMID: 24676717 Free PMC article. Review.
-
A Variant of the Histone-Binding Protein sNASP Contributes to Mouse Lupus.Front Immunol. 2019 Apr 2;10:637. doi: 10.3389/fimmu.2019.00637. eCollection 2019. Front Immunol. 2019. PMID: 31001259 Free PMC article.
-
Roles of histone chaperone Nap1 and histone acetylation in regulating phase-separation of chromatin arrays.bioRxiv [Preprint]. 2025 May 15:2025.05.09.653121. doi: 10.1101/2025.05.09.653121. bioRxiv. 2025. PMID: 40463029 Free PMC article. Preprint.
References
-
- Obuse C, Yang H, Nozaki N, Goto S, Okazaki T, Yoda K. Proteomics analysis of the centromere complex from HeLa interphase cells: UV-damaged DNA binding protein 1 (DDB-1) is a component of the CEN-complex, while BMI-1 is transiently co-localized with the centromeric region in interphase. Genes Cells. 2004;9:105–120. - PubMed
-
- Foltz DR, Jansen LE, Black BE, Bailey AO, Yates JR, III, Cleveland DW. The human CENP-A centromeric nucleosome-associated complex. Nat. Cell Biol. 2006;8:458–469. - PubMed
-
- Okada M, Cheeseman IM, Hori T, Okawa K, McLeod IX, Yates JR, III, Desai A, Fukagawa T. The CENP-H-I complex is required for the efficient incorporation of newly synthesized CENP-A into centromeres. Nat. Cell Biol. 2006;8:446–457. - PubMed
-
- Hori T, Amano M, Suzuki A, Backer CB, Welburn JP, Dong Y, McEwen BF, Shang WH, Suzuki E, Okawa K, et al. CCAN makes multiple contacts with centromeric DNA to provide distinct pathways to the outer kinetochore. Cell. 2008;135:1039–1052. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
