

Id2 influences differentiation of killer cell lectin-like receptor G1(hi) short-lived CD8+ effector T cells
- PMID: 23325888
- PMCID: PMC3563862
- DOI: 10.4049/jimmunol.1200750
Id2 influences differentiation of killer cell lectin-like receptor G1(hi) short-lived CD8+ effector T cells
Abstract
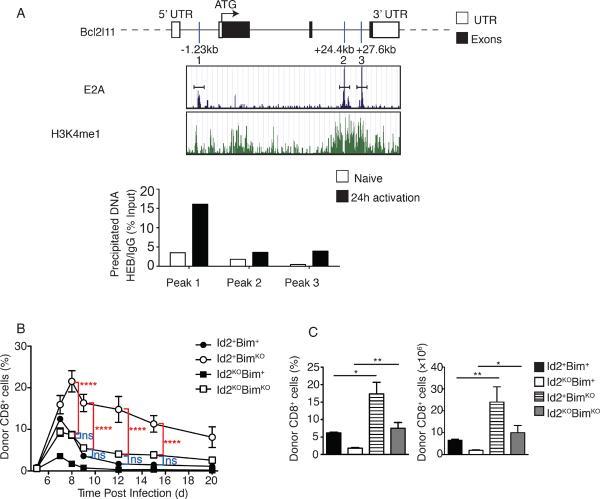
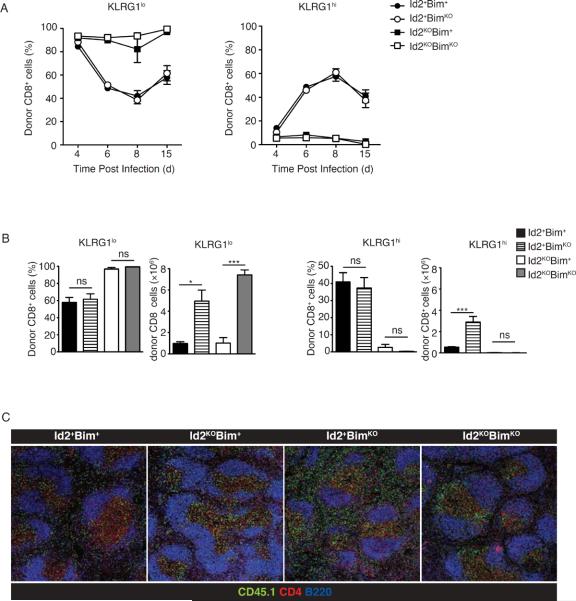
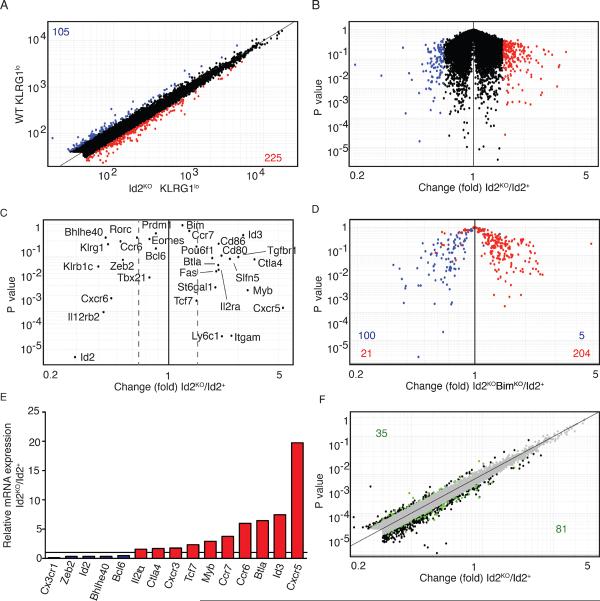
CD8(+) T cells play a crucial role in the clearance of intracellular pathogens through the generation of cytotoxic effector cells that eliminate infected cells and long-lived memory cells that provide enhanced protection against reinfection. We have previously shown that the inhibitor of E protein transcription factors, Id2, is necessary for accumulation of effector and memory CD8(+) T cells during infection. In this study, we show that CD8(+) T cells lacking Id2 did not generate a robust terminally differentiated killer cell lectin-like receptor G1 (KLRG1)(hi) effector population, but displayed a cell-surface phenotype and cytokine profile consistent with memory precursors, raising the question as to whether loss of Id2 impairs the differentiation and/or survival of effector memory cells. We found that deletion of Bim rescued Id2-deficient CD8(+) cell survival during infection. However, the dramatic reduction in KLRG1(hi) cells caused by loss of Id2 remained in the absence of Bim, such that Id2/Bim double-deficient cells form an exclusively KLRG1(lo)CD127(hi) memory precursor population. Thus, we describe a role for Id2 in both the survival and differentiation of normal CD8(+) effector and memory populations.
Figures



Similar articles
-
Bcl-2 allows effector and memory CD8+ T cells to tolerate higher expression of Bim.J Immunol. 2011 May 15;186(10):5729-37. doi: 10.4049/jimmunol.1100102. Epub 2011 Mar 30. J Immunol. 2011. PMID: 21451108 Free PMC article.
-
The transcriptional regulators Id2 and Id3 control the formation of distinct memory CD8+ T cell subsets.Nat Immunol. 2011 Nov 6;12(12):1221-9. doi: 10.1038/ni.2158. Nat Immunol. 2011. PMID: 22057289 Free PMC article.
-
KLRG1+NKG2A+ CD8 T cells mediate protection and participate in memory responses during γ-herpesvirus infection.J Immunol. 2011 Apr 1;186(7):4051-8. doi: 10.4049/jimmunol.1003122. Epub 2011 Feb 23. J Immunol. 2011. PMID: 21346231 Free PMC article.
-
Remembering one's ID/E-ntity: E/ID protein regulation of T cell memory.Curr Opin Immunol. 2013 Oct;25(5):660-6. doi: 10.1016/j.coi.2013.09.004. Epub 2013 Oct 1. Curr Opin Immunol. 2013. PMID: 24094885 Free PMC article. Review.
-
Surviving the crash: transitioning from effector to memory CD8+ T cell.Semin Immunol. 2009 Apr;21(2):92-8. doi: 10.1016/j.smim.2009.02.002. Epub 2009 Mar 6. Semin Immunol. 2009. PMID: 19269192 Free PMC article. Review.
Cited by
-
Id2 and Id3 maintain the regulatory T cell pool to suppress inflammatory disease.Nat Immunol. 2014 Aug;15(8):767-76. doi: 10.1038/ni.2928. Epub 2014 Jun 29. Nat Immunol. 2014. PMID: 24973820 Free PMC article.
-
Signaling networks controlling ID and E protein activity in T cell differentiation and function.Front Immunol. 2022 Aug 2;13:964581. doi: 10.3389/fimmu.2022.964581. eCollection 2022. Front Immunol. 2022. PMID: 35983065 Free PMC article. Review.
-
Sustained Id2 regulation of E proteins is required for terminal differentiation of effector CD8+ T cells.J Exp Med. 2018 Mar 5;215(3):773-783. doi: 10.1084/jem.20171584. Epub 2018 Feb 12. J Exp Med. 2018. PMID: 29440362 Free PMC article.
-
A molecular threshold for effector CD8(+) T cell differentiation controlled by transcription factors Blimp-1 and T-bet.Nat Immunol. 2016 Apr;17(4):422-32. doi: 10.1038/ni.3410. Epub 2016 Mar 7. Nat Immunol. 2016. PMID: 26950239 Free PMC article.
-
T-cell dysfunction during blinatumomab therapy in pediatric acute lymphoblastic leukemia.Blood Adv. 2025 Aug 12;9(15):3689-3693. doi: 10.1182/bloodadvances.2025015894. Blood Adv. 2025. PMID: 40267221 Free PMC article.
References
-
- Harty JT, Badovinac VP. Shaping and reshaping CD8+ T-cell memory. Nat Rev Immunol. 2008;8:107–119. - PubMed
-
- Joshi NS, Kaech SM. Effector CD8 T cell development: a balancing act between memory cell potential and terminal differentiation. J Immunol. 2008;180:1309–1315. - PubMed
-
- Kaech SM, Hemby S, Kersh E, Ahmed R. Molecular and functional profiling of memory CD8 T cell differentiation. Cell. 2002;111:837–851. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
