

Viral and bacterial interactions in the upper respiratory tract
- PMID: 23326226
- PMCID: PMC3542149
- DOI: 10.1371/journal.ppat.1003057
Viral and bacterial interactions in the upper respiratory tract
Abstract
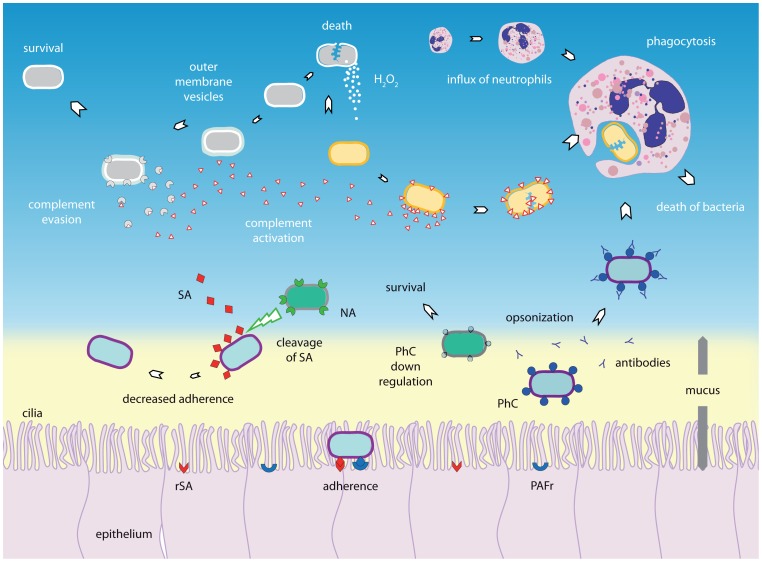
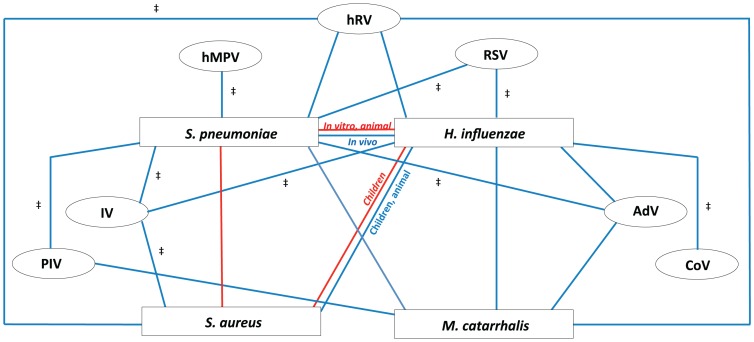
Respiratory infectious diseases are mainly caused by viruses or bacteria that often interact with one another. Although their presence is a prerequisite for subsequent infections, viruses and bacteria may be present in the nasopharynx without causing any respiratory symptoms. The upper respiratory tract hosts a vast range of commensals and potential pathogenic bacteria, which form a complex microbial community. This community is assumed to be constantly subject to synergistic and competitive interspecies interactions. Disturbances in the equilibrium, for instance due to the acquisition of new bacteria or viruses, may lead to overgrowth and invasion. A better understanding of the dynamics between commensals and pathogens in the upper respiratory tract may provide better insight into the pathogenesis of respiratory diseases. Here we review the current knowledge regarding specific bacterial-bacterial and viral-bacterial interactions that occur in the upper respiratory niche, and discuss mechanisms by which these interactions might be mediated. Finally, we propose a theoretical model to summarize and illustrate these mechanisms.
Conflict of interest statement
DB and KT declare to have received consulting fees from Pfizer. EAMS declares to have received unrestricted research support from Pfizer and Baxter, consulting fees from Pfizer and GlaxoSmithKline, lecturing fees from Pfizer, and grant support for vaccine studies from Pfizer and GlaxoSmithKline. None of the fees or grants listed here were received for the research described in this paper. For all other authors no conflicts of interest were declared. This does not alter our adherence to all
Figures

References
-
- World Health Organization Geneva, Health Statistics and Informatics Department (2011) Causes of death 2008. In: World health statistics 2011. 170 pages, ISBN 978 92 4 156419 9 (NLM classification: WA 900.1). pp. 57–76. Available: http://www.who.int/gho/publications/world_health_statistics/EN_WHS2011_F.... Accessed 7 December 2012.
-
- Black RE, Cousens S, Johnson HL, Lawn JE, Rudan I, et al. (2010) Global, regional, and national causes of child mortality in 2008: A systematic analysis. child health epidemiology reference group of WHO and UNICEF. Lancet 375(9730) 1969–1987. - PubMed
-
- Vergison A, Dagan R, Arguedas A, Bonhoeffer J, Cohen R, et al. (2010) Otitis media and its consequences: Beyond the earache. Lancet Infect Dis 10(3) 195–203 10.1016/S1473-3099(10)70012-8. - PubMed
-
- Watson KBH, Carville KMA, Bowman JBA, Jacoby P, Riley TV, et al. (2006) Upper respiratory tract bacterial carriage in aboriginal and non-aboriginal children in a semi-arid area of western Australia. Pediatr Infect Dis J 25(9) 782–790 10.1097/01.inf.0000232705.49634.68. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
