

Pre-mRNA splicing is a determinant of nucleosome organization
- PMID: 23326444
- PMCID: PMC3542351
- DOI: 10.1371/journal.pone.0053506
Pre-mRNA splicing is a determinant of nucleosome organization
Abstract
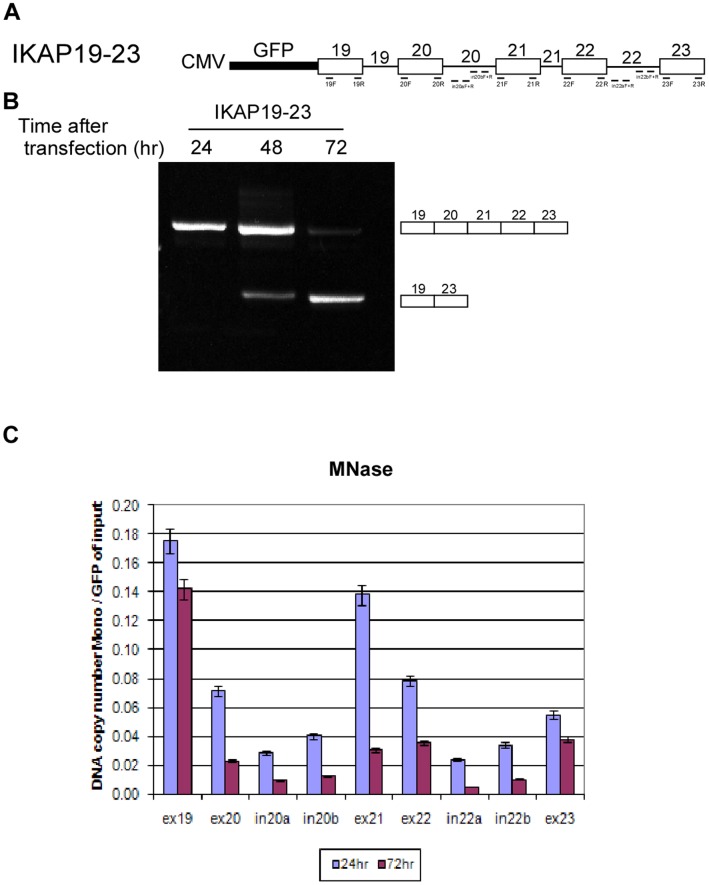
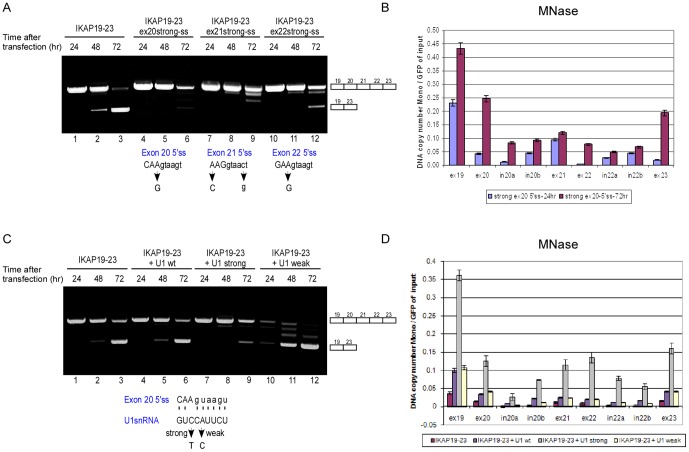
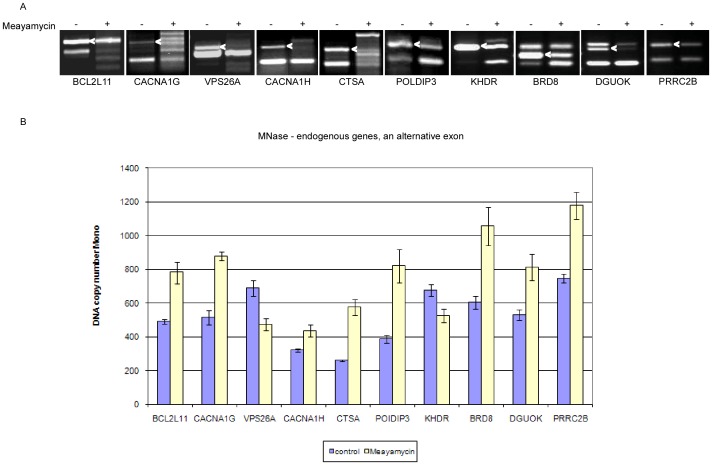
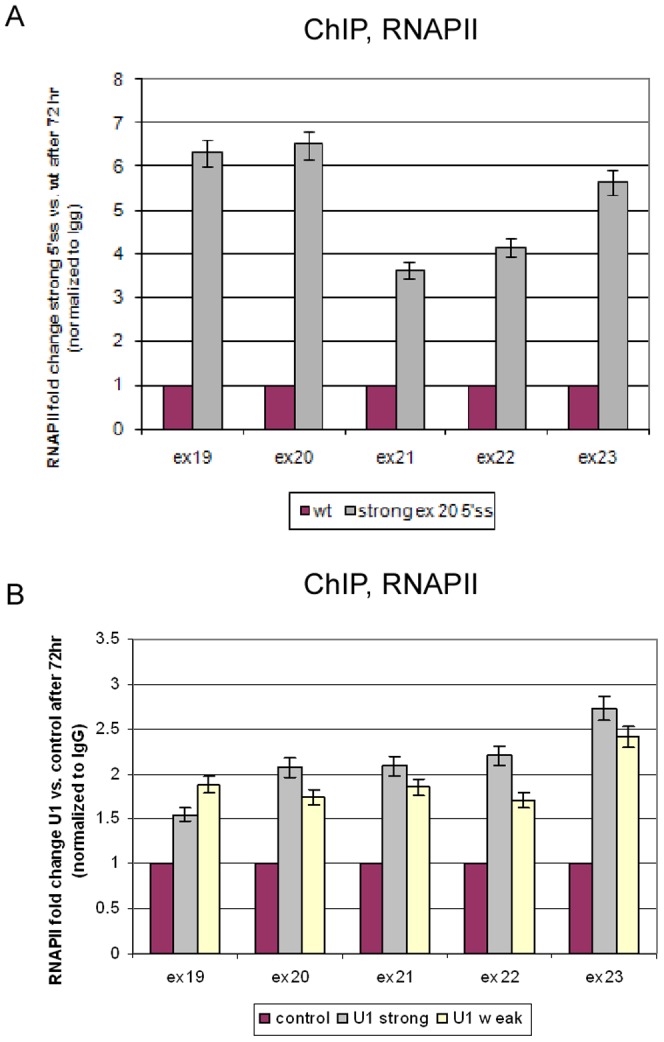
Chromatin organization affects alternative splicing and previous studies have shown that exons have increased nucleosome occupancy compared with their flanking introns. To determine whether alternative splicing affects chromatin organization we developed a system in which the alternative splicing pattern switched from inclusion to skipping as a function of time. Changes in nucleosome occupancy were correlated with the change in the splicing pattern. Surprisingly, strengthening of the 5' splice site or strengthening the base pairing of U1 snRNA with an internal exon abrogated the skipping of the internal exons and also affected chromatin organization. Over-expression of splicing regulatory proteins also affected the splicing pattern and changed nucleosome occupancy. A specific splicing inhibitor was used to show that splicing impacts nucleosome organization endogenously. The effect of splicing on the chromatin required a functional U1 snRNA base pairing with the 5' splice site, but U1 pairing was not essential for U1 snRNA enhancement of transcription. Overall, these results suggest that splicing can affect chromatin organization.
Conflict of interest statement
Figures
Similar articles
-
Chromatin's thread to alternative splicing regulation.Chromosoma. 2013 Dec;122(6):465-74. doi: 10.1007/s00412-013-0425-x. Epub 2013 Aug 3. Chromosoma. 2013. PMID: 23912688 Review.
-
Relationship between nucleosome positioning and progesterone-induced alternative splicing in breast cancer cells.RNA. 2015 Mar;21(3):360-74. doi: 10.1261/rna.048843.114. RNA. 2015. PMID: 25589247 Free PMC article.
-
Differential patterns of intronic and exonic DNA regions with respect to RNA polymerase II occupancy, nucleosome density and H3K36me3 marking in fission yeast.Genome Biol. 2011 Aug 22;12(8):R82. doi: 10.1186/gb-2011-12-8-r82. Genome Biol. 2011. PMID: 21859475 Free PMC article.
-
Extended base pair complementarity between U1 snRNA and the 5' splice site does not inhibit splicing in higher eukaryotes, but rather increases 5' splice site recognition.Nucleic Acids Res. 2005 Sep 9;33(16):5112-9. doi: 10.1093/nar/gki824. Print 2005. Nucleic Acids Res. 2005. PMID: 16155183 Free PMC article.
-
Role and mechanism of U1-independent pre-mRNA splicing in the regulation of alternative splicing.RNA Biol. 2009 Sep-Oct;6(4):395-8. doi: 10.4161/rna.6.4.9318. Epub 2009 Sep 21. RNA Biol. 2009. PMID: 19667760 Review.
Cited by
-
Insulators recruit histone methyltransferase dMes4 to regulate chromatin of flanking genes.EMBO J. 2014 Jul 17;33(14):1599-613. doi: 10.15252/embj.201385965. Epub 2014 Jun 10. EMBO J. 2014. PMID: 24916307 Free PMC article.
-
The determinants of alternative RNA splicing in human cells.Mol Genet Genomics. 2017 Dec;292(6):1175-1195. doi: 10.1007/s00438-017-1350-0. Epub 2017 Jul 13. Mol Genet Genomics. 2017. PMID: 28707092 Review.
-
Alternative splicing regulates the expression of G9A and SUV39H2 methyltransferases, and dramatically changes SUV39H2 functions.Nucleic Acids Res. 2015 Feb 18;43(3):1869-82. doi: 10.1093/nar/gkv013. Epub 2015 Jan 20. Nucleic Acids Res. 2015. PMID: 25605796 Free PMC article.
-
Chromatin's thread to alternative splicing regulation.Chromosoma. 2013 Dec;122(6):465-74. doi: 10.1007/s00412-013-0425-x. Epub 2013 Aug 3. Chromosoma. 2013. PMID: 23912688 Review.
-
PRPF6 promotes androgen receptor/androgen receptor-variant 7 actions in castration-resistant prostate cancer cells.Int J Biol Sci. 2021 Jan 1;17(1):188-203. doi: 10.7150/ijbs.50810. eCollection 2021. Int J Biol Sci. 2021. PMID: 33390843 Free PMC article.
References
-
- Keren H, Lev-Maor G, Ast G (2010) Alternative splicing and evolution: diversification, exon definition and function. Nat Rev Genet 11: 345–355. - PubMed
-
- de Almeida SF, Carmo-Fonseca M (2012) Design principles of interconnections between chromatin and pre-mRNA splicing. Trends Biochem Sci - PubMed
-
- Alexander R, Beggs JD (2010) Cross-talk in transcription, splicing and chromatin: who makes the first call? Biochem Soc Trans 38: 1251–1256. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
