

Duplication and diversification of the spermidine/spermine N1-acetyltransferase 1 genes in zebrafish
- PMID: 23326562
- PMCID: PMC3543422
- DOI: 10.1371/journal.pone.0054017
Duplication and diversification of the spermidine/spermine N1-acetyltransferase 1 genes in zebrafish
Abstract
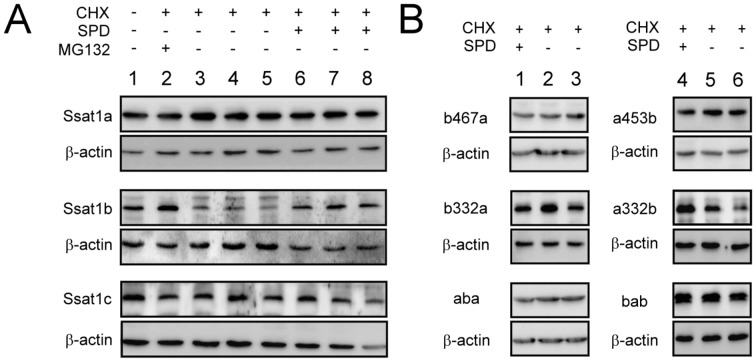
Spermidine/spermine N(1)-acetyltransferase 1 (Ssat1) is a key enzyme in the polyamine interconversion pathway, which maintains polyamine homeostasis. In addition, mammalian Ssat1 is also involved in many physiological and pathological events such as hypoxia, cell migration, and carcinogenesis. Using cross-genomic bioinformatic analysis in 10 deuterostomes, we found that ssat1 only exists in vertebrates. Comparing with mammalian, zebrafish, an evolutionarily distant vertebrate, contains 3 homologous ssat1 genes, named ssat1a, ssat1b, and ssat1c. All zebrafish homologues could be transcribed and produce active enzymes. Despite the long history since their evolutionary diversification, some features of human SSAT1 are conserved and subfunctionalized in the zebrafish family of Ssat1 proteins. The polyamine-dependent protein synthesis was only found in Ssat1b and Ssat1c, not in Ssat1a. Further study indicated that both 5' and 3' sequences of ssat1b mediate such kind of translational regulation inside the open reading frame (ORF). The polyamine-dependent protein stabilization was only observed in Ssat1b. The last 70 residues of Ssat1b were crucial for its rapid degradation and polyamine-induced stabilization. It is worth noting that only Ssat1b and Ssat1c, but not the polyamine-insensitive Ssat1a, were able to interact with integrin α9 and Hif-1α. Thus, Ssat1b and Ssat1c might not only be a polyamine metabolic enzyme but also simultaneously respond to polyamine levels and engage in cross-talk with other signaling pathways. Our data revealed some correlations between the sequences and functions of the zebrafish family of Ssat1 proteins, which may provide valuable information for studies of their translational regulatory mechanism, protein stability, and physiological functions.
Conflict of interest statement
Figures





References
-
- Agostinelli E (2012) Role of polyamines, their analogs and transglutaminases in biological and clinical perspectives. Amino Acids 42: 397–409. - PubMed
-
- Perez-Leal O, Merali S (2012) Regulation of polyamine metabolism by translational control. Amino Acids 42: 611–617. - PubMed
-
- Pegg AE (2008) Spermidine/spermine-N(1)-acetyltransferase: a key metabolic regulator. Am J Physiol Endocrinol Metab 294: E995–1010. - PubMed
-
- Agostinelli E, Tempera G, Viceconte N, Saccoccio S, Battaglia V, et al. (2010) Potential anticancer application of polyamine oxidation products formed by amine oxidase: a new therapeutic approach. Amino Acids 38: 353–368. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
