

ATG5 regulates plasma cell differentiation
- PMID: 23327930
- PMCID: PMC3627668
- DOI: 10.4161/auto.23484
ATG5 regulates plasma cell differentiation
Abstract
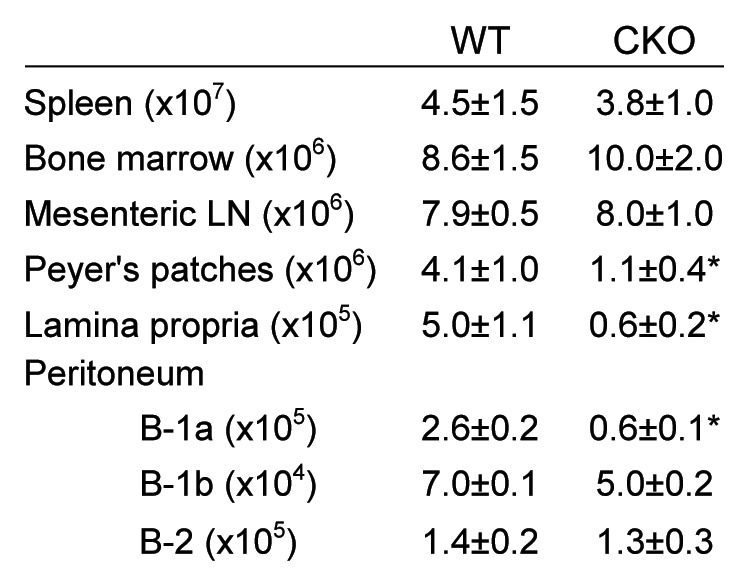
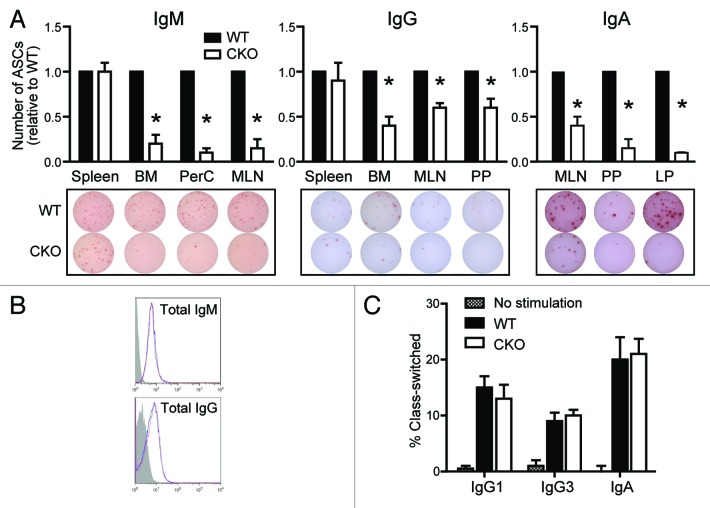
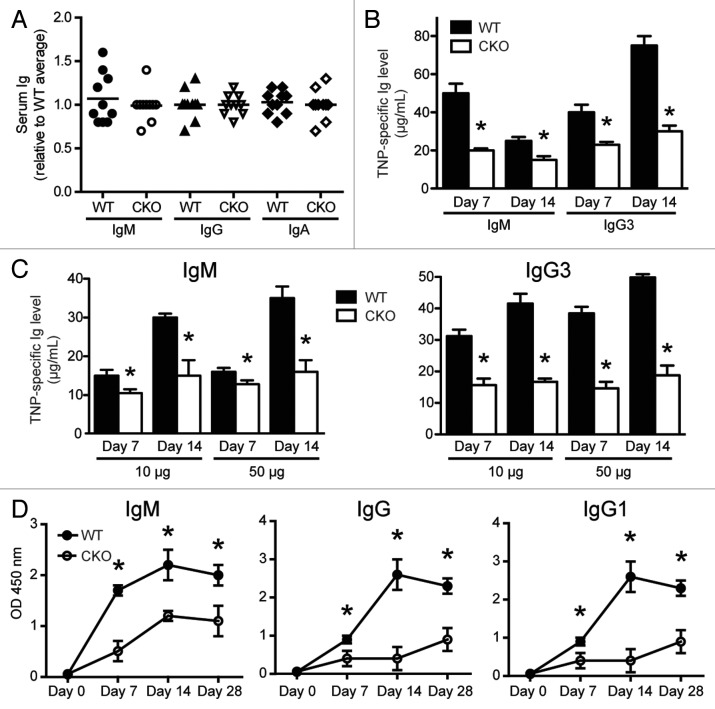
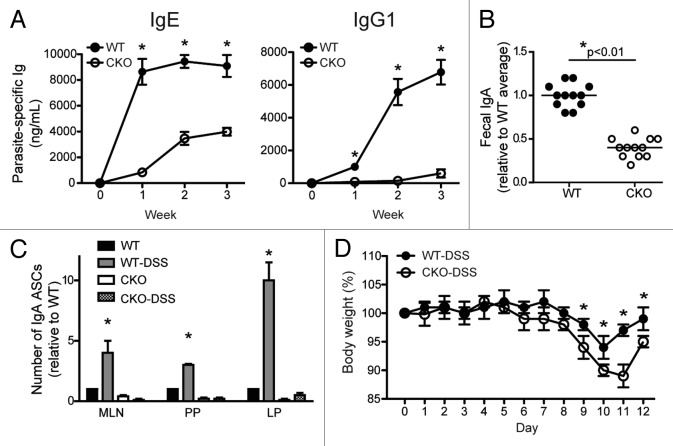
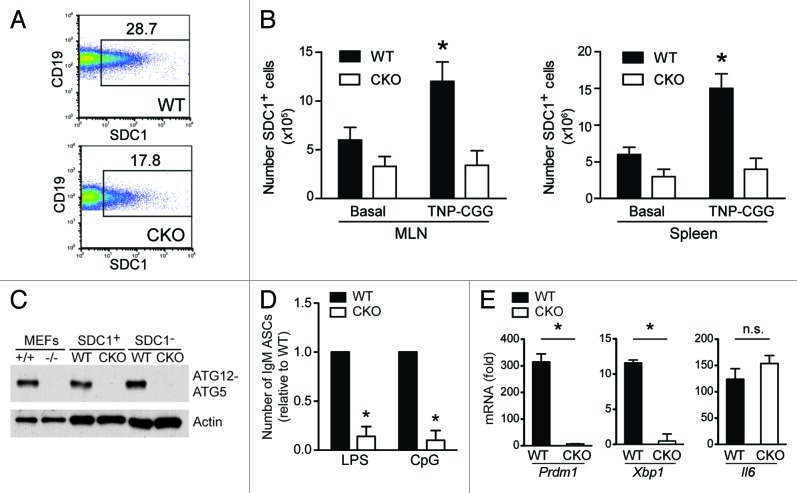
Autophagy is a conserved homeostatic process in which cytoplasmic contents are degraded and recycled. Two ubiquitin-like conjugation pathways are required for the generation of autophagosomes, and ATG5 is necessary for both of these processes. Studies of mice deficient in ATG5 reveal a key role for autophagy in T lymphocyte function, as well as in B cell development and B-1a B cell maintenance. However, the role of autophagy genes in B cell function and antibody production has not been described. Using mice in which Atg5 is conditionally deleted in B lymphocytes, we showed here that this autophagy gene is essential for plasma cell homeostasis. In the absence of B cell ATG5 expression, antibody responses were significantly diminished during antigen-specific immunization, parasitic infection and mucosal inflammation. Atg5-deficient B cells maintained the ability to produce immunoglobulin and undergo class-switch recombination, yet had impaired SDC1 expression, significantly decreased antibody secretion in response to toll-like receptor ligands, and an inability to upregulate plasma cell transcription factors. These results build upon previous data demonstrating a role for ATG5 in early B cell development, illustrating its importance in late B cell activation and subsequent plasma cell differentiation.
Keywords: ATG5; B lymphocytes; antibody secretion; immunity; plasma cell differentiation.
Figures
Similar articles
-
Plasma cells require autophagy for sustainable immunoglobulin production.Nat Immunol. 2013 Mar;14(3):298-305. doi: 10.1038/ni.2524. Epub 2013 Jan 27. Nat Immunol. 2013. PMID: 23354484
-
A critical role for the autophagy gene Atg5 in T cell survival and proliferation.J Exp Med. 2007 Jan 22;204(1):25-31. doi: 10.1084/jem.20061303. Epub 2006 Dec 26. J Exp Med. 2007. PMID: 17190837 Free PMC article.
-
CD36 and LC3B initiated autophagy in B cells regulates the humoral immune response.Autophagy. 2021 Nov;17(11):3577-3591. doi: 10.1080/15548627.2021.1885183. Epub 2021 Feb 16. Autophagy. 2021. PMID: 33535890 Free PMC article.
-
Autophagy, a new determinant of plasma cell differentiation and antibody responses.Mol Immunol. 2014 Dec;62(2):289-95. doi: 10.1016/j.molimm.2014.02.008. Epub 2014 Mar 11. Mol Immunol. 2014. PMID: 24629228 Review.
-
Re-thinking the functions of IgA(+) plasma cells.Gut Microbes. 2014;5(5):652-62. doi: 10.4161/19490976.2014.969977. Gut Microbes. 2014. PMID: 25483334 Free PMC article. Review.
Cited by
-
Autophagy: roles in intestinal mucosal homeostasis and inflammation.J Biomed Sci. 2019 Feb 14;26(1):19. doi: 10.1186/s12929-019-0512-2. J Biomed Sci. 2019. PMID: 30764829 Free PMC article. Review.
-
ATRA-induced NEAT1 upregulation promotes autophagy during APL cell granulocytic differentiation.PLoS One. 2024 Dec 23;19(12):e0316109. doi: 10.1371/journal.pone.0316109. eCollection 2024. PLoS One. 2024. PMID: 39715205 Free PMC article.
-
The Mucosal Immune System and Its Regulation by Autophagy.Front Immunol. 2016 Jun 22;7:240. doi: 10.3389/fimmu.2016.00240. eCollection 2016. Front Immunol. 2016. PMID: 27446072 Free PMC article. Review.
-
Small Molecule Inhibition of Rab7 Impairs B Cell Class Switching and Plasma Cell Survival To Dampen the Autoantibody Response in Murine Lupus.J Immunol. 2016 Nov 15;197(10):3792-3805. doi: 10.4049/jimmunol.1601427. Epub 2016 Oct 14. J Immunol. 2016. PMID: 27742832 Free PMC article.
-
Autophagy and the Immune Response.Adv Exp Med Biol. 2019;1206:595-634. doi: 10.1007/978-981-15-0602-4_27. Adv Exp Med Biol. 2019. PMID: 31777004 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous