

Review
doi: 10.1038/nrg3366.
Y-chromosome evolution: emerging insights into processes of Y-chromosome degeneration
Affiliations
- PMID: 23329112
- PMCID: PMC4120474
- DOI: 10.1038/nrg3366
Item in Clipboard
Review
Y-chromosome evolution: emerging insights into processes of Y-chromosome degeneration
Nat Rev Genet.
2013 Feb.
Abstract
The human Y chromosome is intriguing not only because it harbours the master-switch gene that determines gender but also because of its unusual evolutionary history. The Y chromosome evolved from an autosome, and its evolution has been characterized by massive gene decay. Recent whole-genome and transcriptome analyses of Y chromosomes in humans and other primates, in Drosophila species and in plants have shed light on the current gene content of the Y chromosome, its origins and its long-term fate. Furthermore, comparative analysis of young and old Y chromosomes has given further insights into the evolutionary and molecular forces triggering Y-chromosome degeneration and into the evolutionary destiny of the Y chromosome.
Figures
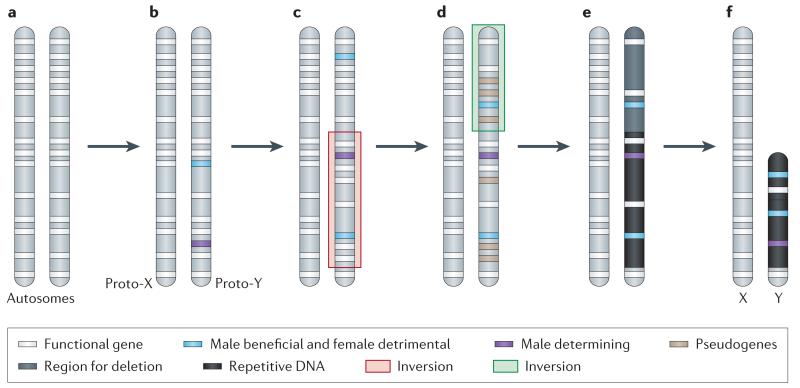
a. Sex chromosomes formed from ordinary autosomes, containing identical sets of genes b. A potential first step in the evolution of heteromorphic sex chromosomes is the acquisition of a sex-determining locus on a proto-sex chromosome, such as a male-determining gene. The emergence of separate sexes and sex chromosomes from a hermaphrodite ancestor requires a male- and a female-sterility mutation to occur on the proto-sex chromosome. c. Accumulation of sexually antagonistic mutations close to the sex-determining region select for suppression of recombination on the proto-sex chromosomes which can be achieved by chromosomal inversion. d. The non-recombining region can increase, if other mutations with sex-specific fitness effects accumulate on proto-sex chromosomes. Furthermore, a lack of recombination results in the accumulation of loss-of function mutations at Y-linked genes (pseudogenization). e. Lack of recombination also results in an accumulation of repetitive DNA, which can lead to an increase of the size of the evolving Y. f. Large segments of non-functional DNA can be deleted in old Y chromosomes, and reduce their physical size. The evolutionary outcome of this process are heteromorphic sex chromosomes, where the X chromosome largely resembles the autosome from which it derived, and the Y chromosomes has lost most of its ancestral genes, and may instead have accumulated repetitive DNA.
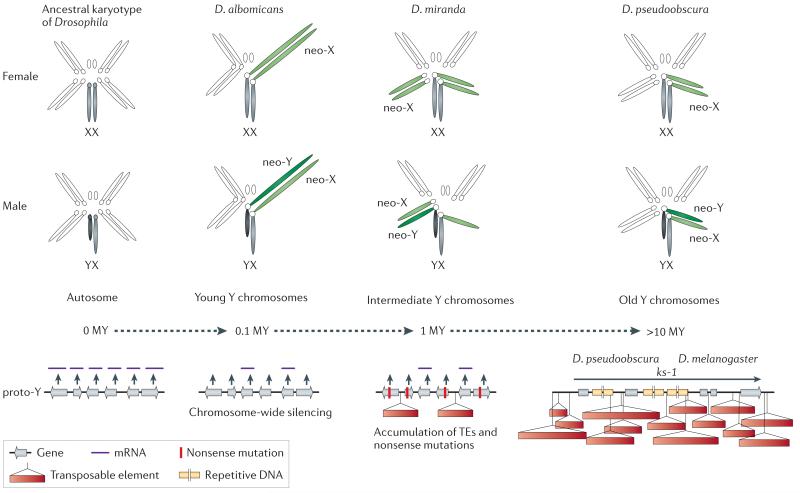
Neo-sex chromosomes (in green) are formed by fusions of autosomes with the ancestral sex chromosomes (in grey), and the neo-X and neo-Y carry identical gene sets at the time of their origination (0 MY). To date, three Drosophila species with neo-Y’s of different age have been studied in detail, and provide a dynamic picture of the molecular changes associated with Y differentiation, and their karyotype is shown. Chromosome-wide down-regulation of protein-coding genes before accumulation of nonsense mutations at coding sequences is observed on the neo-Y in D. albomicans, which originated about 0.1 MY ago. Thus, gene decay on the neo-Y appears to be initiated by chromosome-wide silencing, possibly as a result of epigenetic modifications causing a change in chromatin structure. A general alteration of the genome architecture of the neo-Y has occurred within only 1 MY in D. miranda. In particular, the neo-Y of this species is characterized by massive decay in gene function, including the fixation of stop codons and frame shift mutations, large deletions and a general accumulation of repetitive transposable element DNA. The loss of gene function is accompanied by the beginning of heterochromatinization of the neo-Y. After 15 MY, almost no sequence similarity between the neo-Y and its former homolog can be detected, and the neo-Y has become entirely heterochromatic. This evolutionary stage has been reached in D. pseudoobscura, and resembles the general architecture of the ancestral Y in Drosophila. Genes on the ancestral Y of Drosophila contain massive introns, filled with repetitive DNA. Y-linked genes in Drosophila, such as ks-1, are characterized by huge introns, filled with repetitive satellite DNA.

a. Population genetic processes to explain gene loss on a non-recombining chromosome do not predict continuous gene decay and eventual extinction of the Y,. A simple model of constant gene decay is shown by the red dashed line. Instead, analytical approximations (green line) and computer simulations (asterisk) under the Muller’s ratchet and genetic hitchhiking model suggest that gene-rich non-recombining chromosomes degenerate rapidly initially, but gene decay slows down over time and ultimately comes to a complete halt once a threshold number of Y-linked genes has been reached. In addition, empirical evidence from Drosophila neo-Y chromosome sequencing and primate Y chromosome gene content are consistent with this non-linear degeneration.
References
-
- Lahn BT, Page DC. Four evolutionary strata on the human X chromosome. Science. 1999;286:964–967. - PubMed
-
- Charlesworth B. The evolution of sex chromosomes. Science. 1990;251:1030–1033. - PubMed
-
- Bull JJ. Evolution of Sex Determining Mechanisms. Benjamin Cummings; Menlo Park, CA: 1983.
-
- Rice WR. Evolution of the Y sex chromosome in animals. BioScience. 1996;46:331–343.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
