

The cotranslational function of ribosome-associated Hsp70 in eukaryotic protein homeostasis
- PMID: 23332755
- PMCID: PMC3553497
- DOI: 10.1016/j.cell.2012.12.001
The cotranslational function of ribosome-associated Hsp70 in eukaryotic protein homeostasis
Abstract
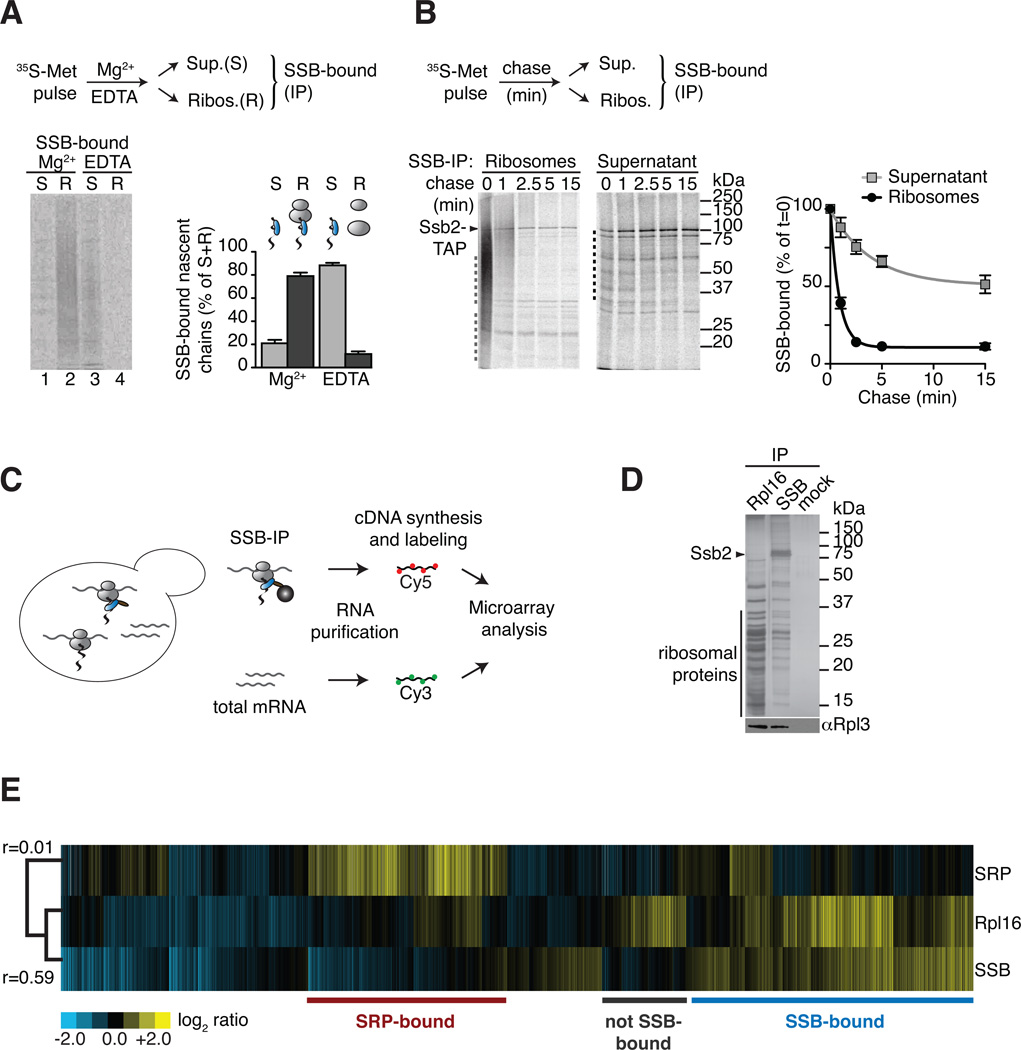
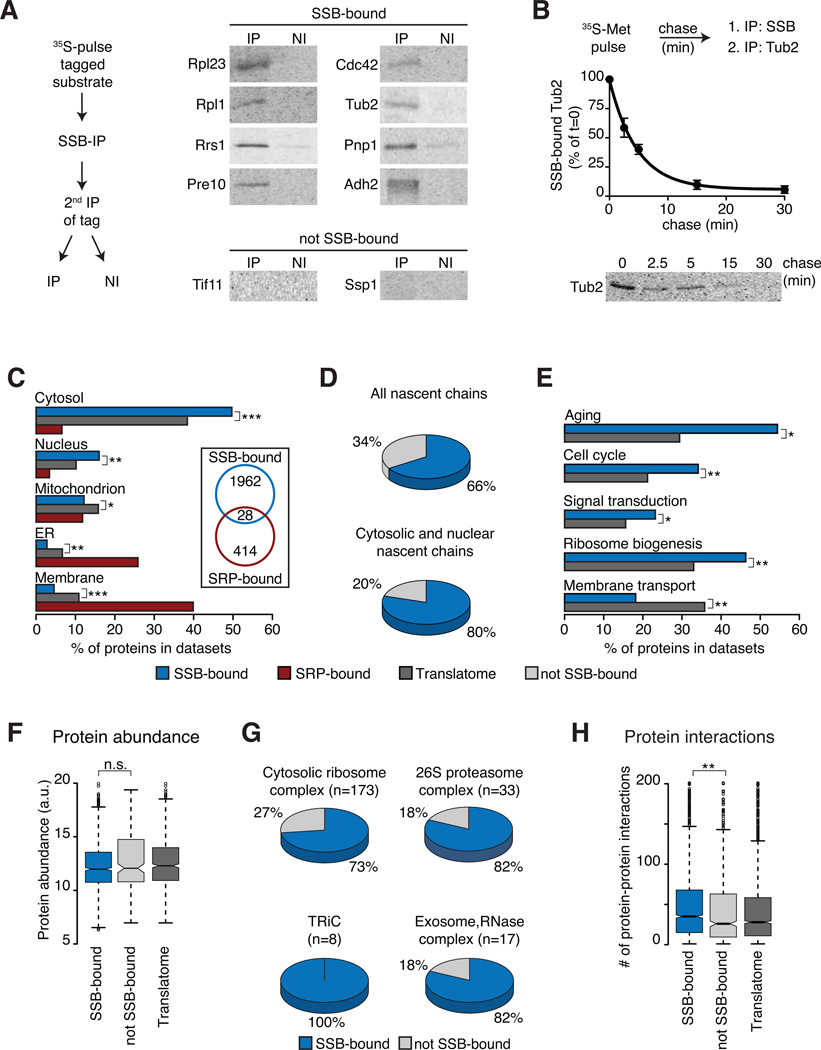
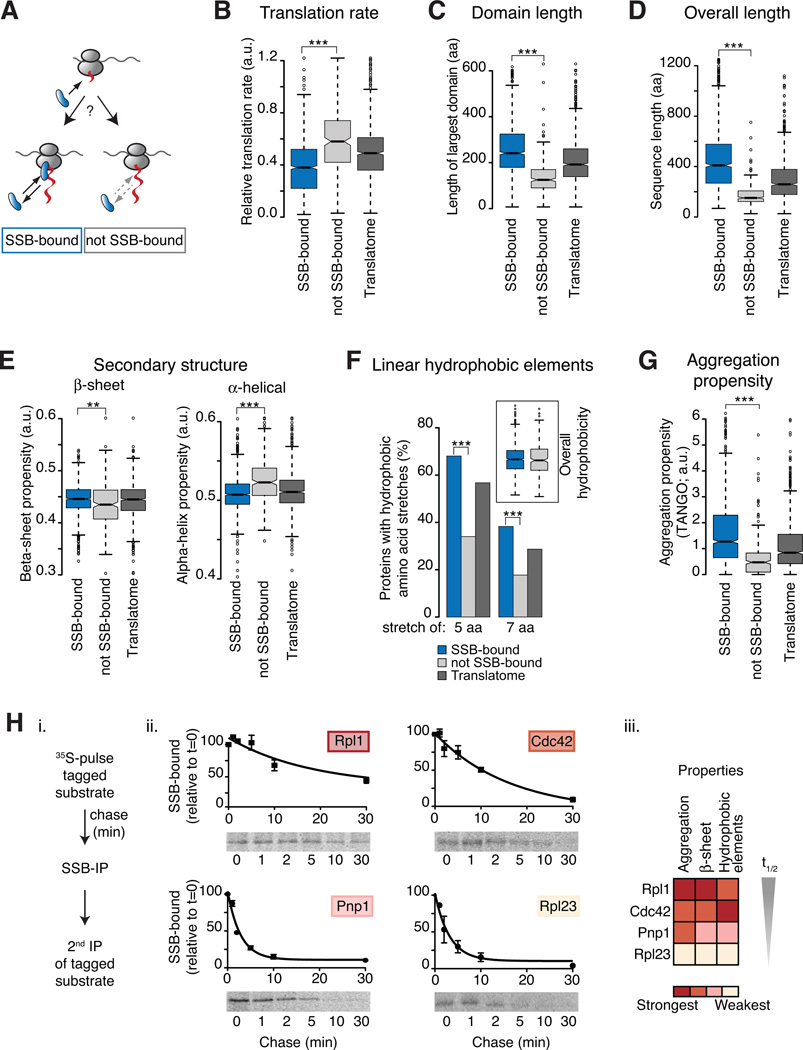
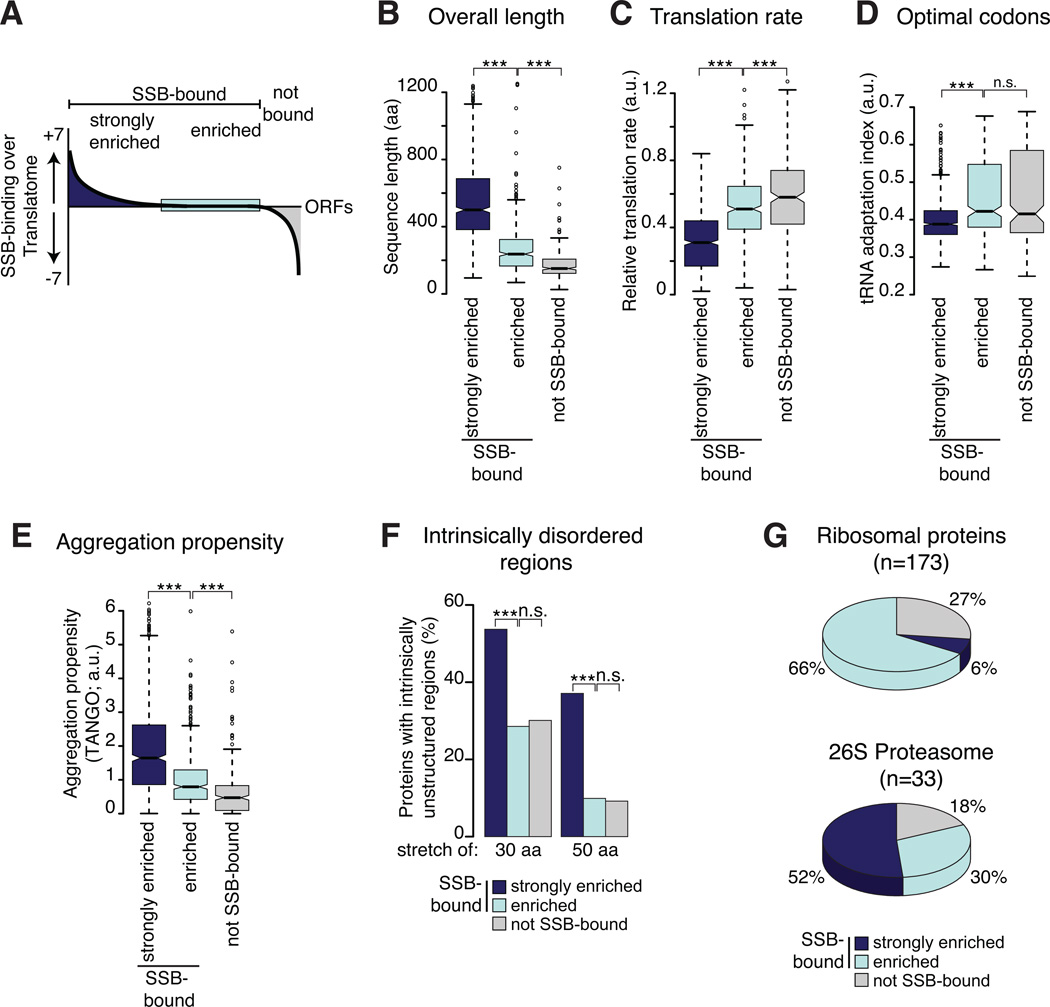
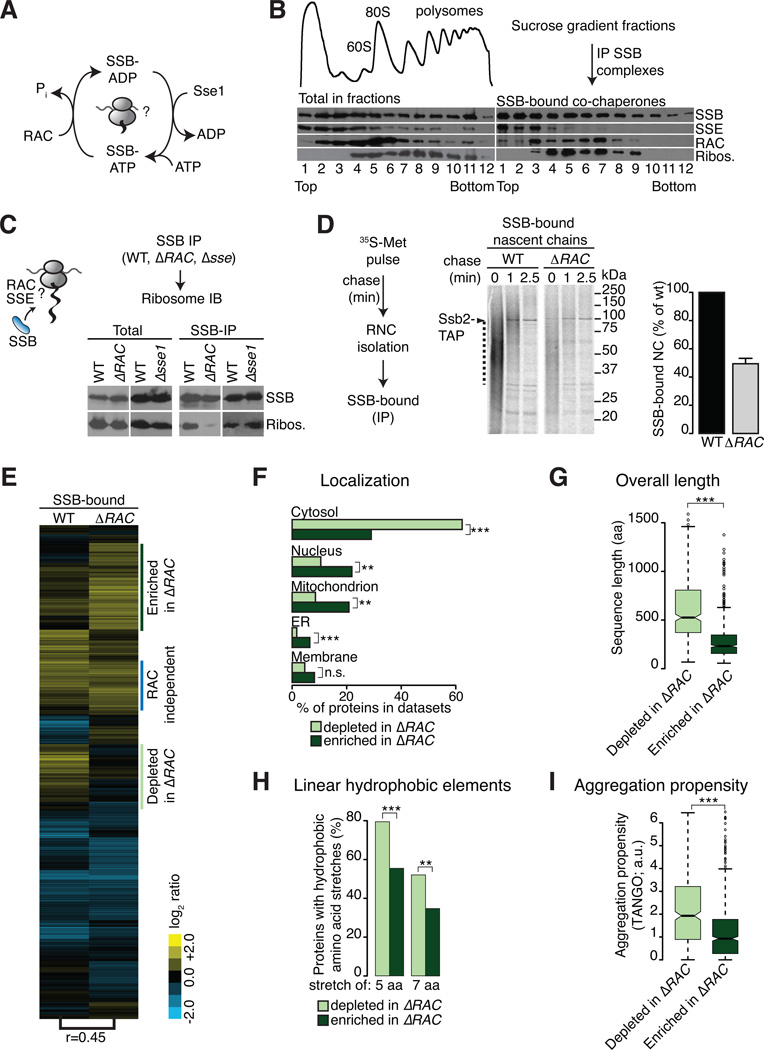
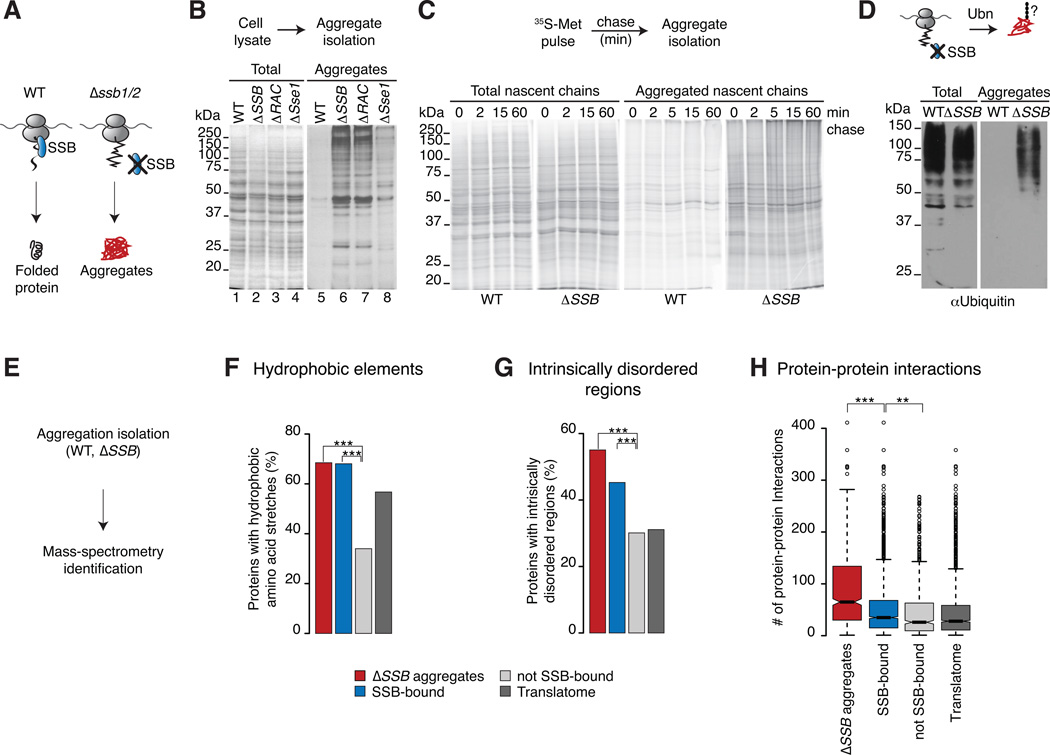
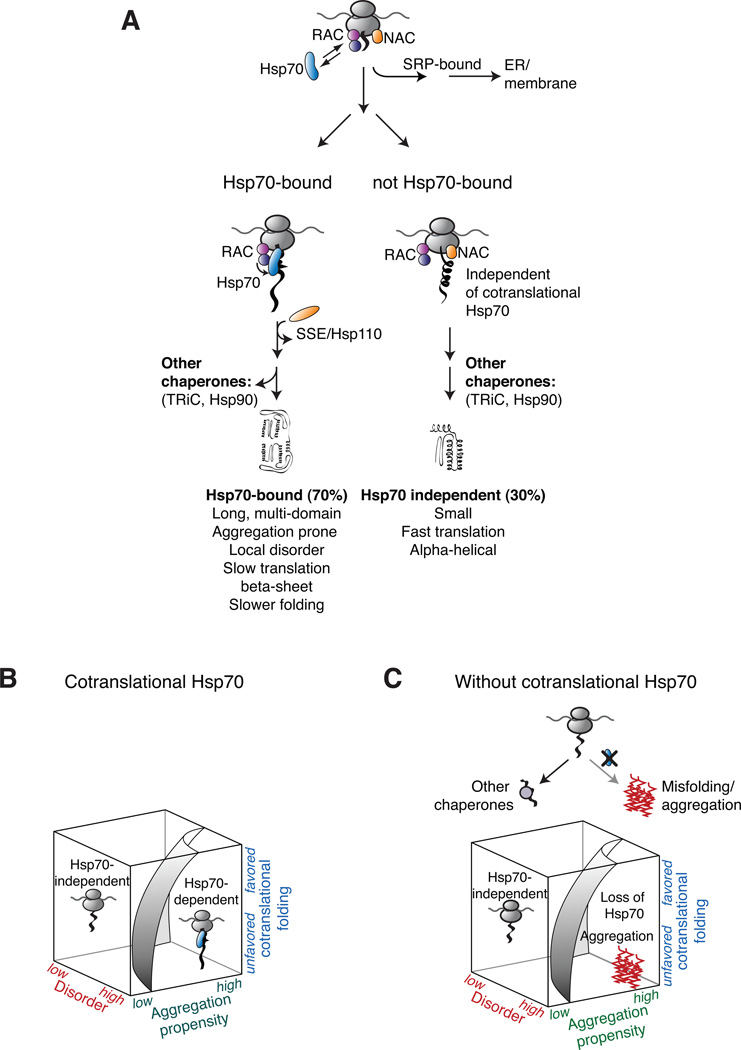
In eukaryotic cells a molecular chaperone network associates with translating ribosomes, assisting the maturation of emerging nascent polypeptides. Hsp70 is perhaps the major eukaryotic ribosome-associated chaperone and the first reported to bind cotranslationally to nascent chains. However, little is known about the underlying principles and function of this interaction. Here, we use a sensitive and global approach to define the cotranslational substrate specificity of the yeast Hsp70 SSB. We find that SSB binds to a subset of nascent polypeptides whose intrinsic properties and slow translation rates hinder efficient cotranslational folding. The SSB-ribosome cycle and substrate recognition is modulated by its ribosome-bound cochaperone, RAC. Deletion of SSB leads to widespread aggregation of newly synthesized polypeptides. Thus, cotranslationally acting Hsp70 meets the challenge of folding the eukaryotic proteome by stabilizing its longer, more slowly translated, and aggregation-prone nascent polypeptides.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Agashe VR, Guha S, Chang H-C, Genevaux P, Hayer-Hartl M, Stemp M, Georgopoulos C, Hartl FU, Barral JM. Function of trigger factor and DnaK in multidomain protein folding: increase in yield at the expense of folding speed. Cell. 2004;117:199–209. - PubMed
-
- Albanese V, Yam AY-W, Baughman J, Parnot C, Frydman J. Systems Analyses Reveal Two Chaperone Networks with Distinct Functions in Eukaryotic Cells. Cell. 2006;124:75–88. - PubMed
-
- Beckmann RP, Mizzen LE, Welch WJ. Interaction of Hsp 70 with newly synthesized proteins: implications for protein folding and assembly. Science. 1990;248:850–854. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
