

An Eco1-independent sister chromatid cohesion establishment pathway in S. cerevisiae
- PMID: 23334284
- PMCID: PMC3608886
- DOI: 10.1007/s00412-013-0396-y
An Eco1-independent sister chromatid cohesion establishment pathway in S. cerevisiae
Abstract
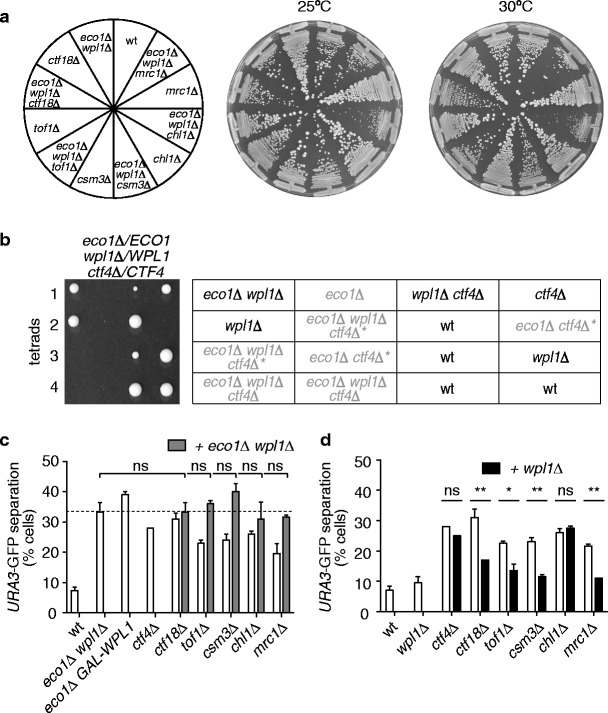
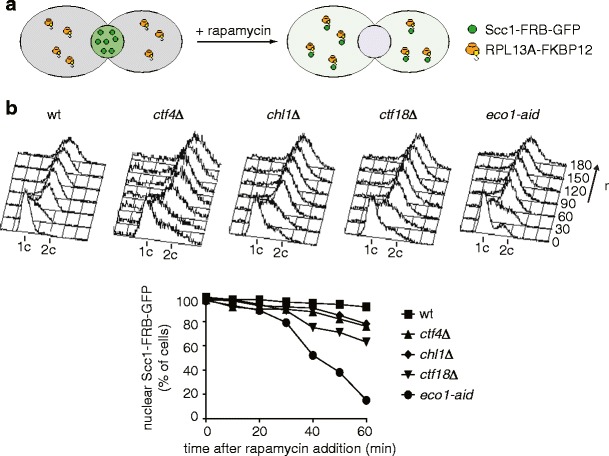
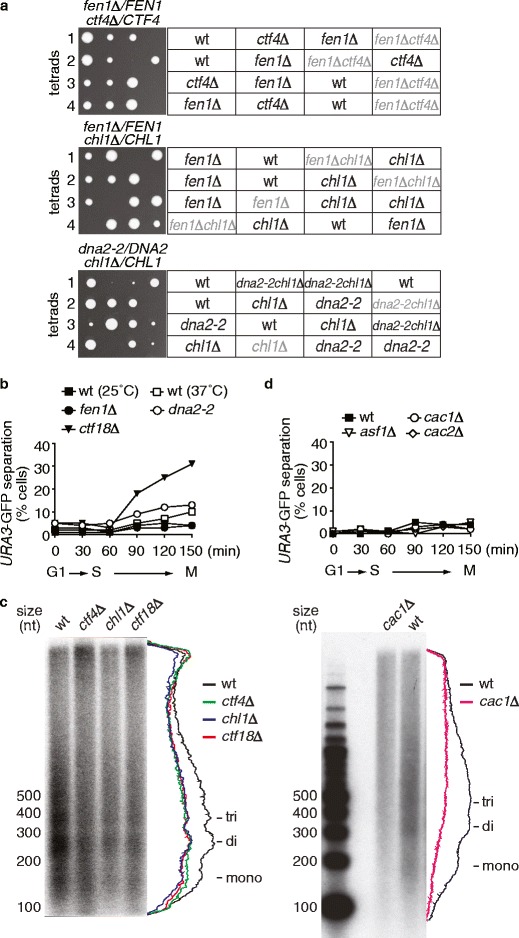
Cohesion between sister chromatids, mediated by the chromosomal cohesin complex, is a prerequisite for their alignment on the spindle apparatus and segregation in mitosis. Budding yeast cohesin first associates with chromosomes in G1. Then, during DNA replication in S-phase, the replication fork-associated acetyltransferase Eco1 acetylates the cohesin subunit Smc3 to make cohesin's DNA binding resistant to destabilization by the Wapl protein. Whether stabilization of cohesin molecules that happen to link sister chromatids is sufficient to build sister chromatid cohesion, or whether additional reactions are required to establish these links, is not known. In addition to Eco1, several other factors contribute to cohesion establishment, including Ctf4, Ctf18, Tof1, Csm3, Chl1 and Mrc1, but little is known about their roles. Here, we show that each of these factors facilitates cohesin acetylation. Moreover, the absence of Ctf4 and Chl1, but not of the other factors, causes a synthetic growth defect in cells lacking Eco1. Distinct from acetylation defects, sister chromatid cohesion in ctf4Δ and chl1Δ cells is not improved by removing Wapl. Unlike previously thought, we do not find evidence for a role of Ctf4 and Chl1 in Okazaki fragment processing, or of Okazaki fragment processing in sister chromatid cohesion. Thus, Ctf4 and Chl1 delineate an additional acetylation-independent pathway that might hold important clues as to the mechanism of sister chromatid cohesion establishment.
Figures



References
-
- Bermudez VP, Maniwa Y, Tappin I, Ozato K, Yokomori K, Hurwitz J. The alternative Ctf18–Dcc1–Ctf8-replication factor C complex required for sister chromatid cohesion loads proliferating cell nuclear antigen onto DNA. Proc Natl Acad Sci USA. 2003;100:10237–10242. doi: 10.1073/pnas.1434308100. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
