

PDGF receptor alpha+ mesoderm contributes to endothelial and hematopoietic cells in mice
- PMID: 23335233
- PMCID: PMC3597973
- DOI: 10.1002/dvdy.23923
PDGF receptor alpha+ mesoderm contributes to endothelial and hematopoietic cells in mice
Abstract
Background: Early mesoderm can be classified into Flk-1+ or PDGF receptor alpha (PDGFRα)+ population, grossly representing lateral and paraxial mesoderm, respectively. It has been demonstrated that all endothelial (EC) and hematopoietic (HPC) cells are derived from Flk-1+ cells. Although PDGFRα+ cells give rise to ECs/HPCs in in vitro ES differentiation, whether PDGFRα+ population can become hemato-endothelial lineages has not been proved in mouse embryos.
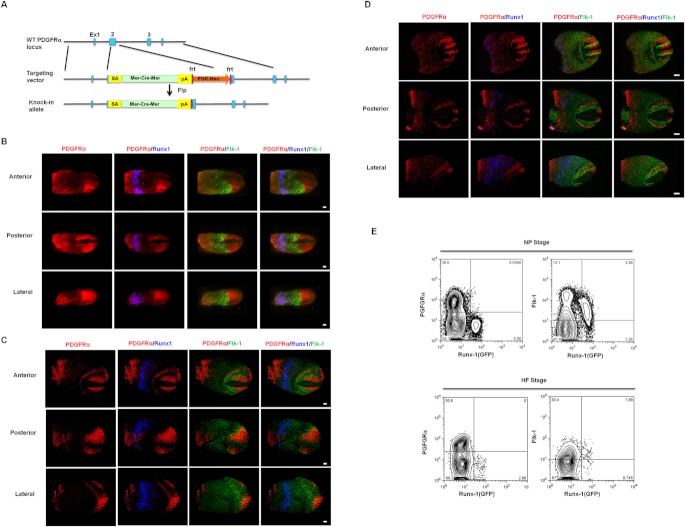
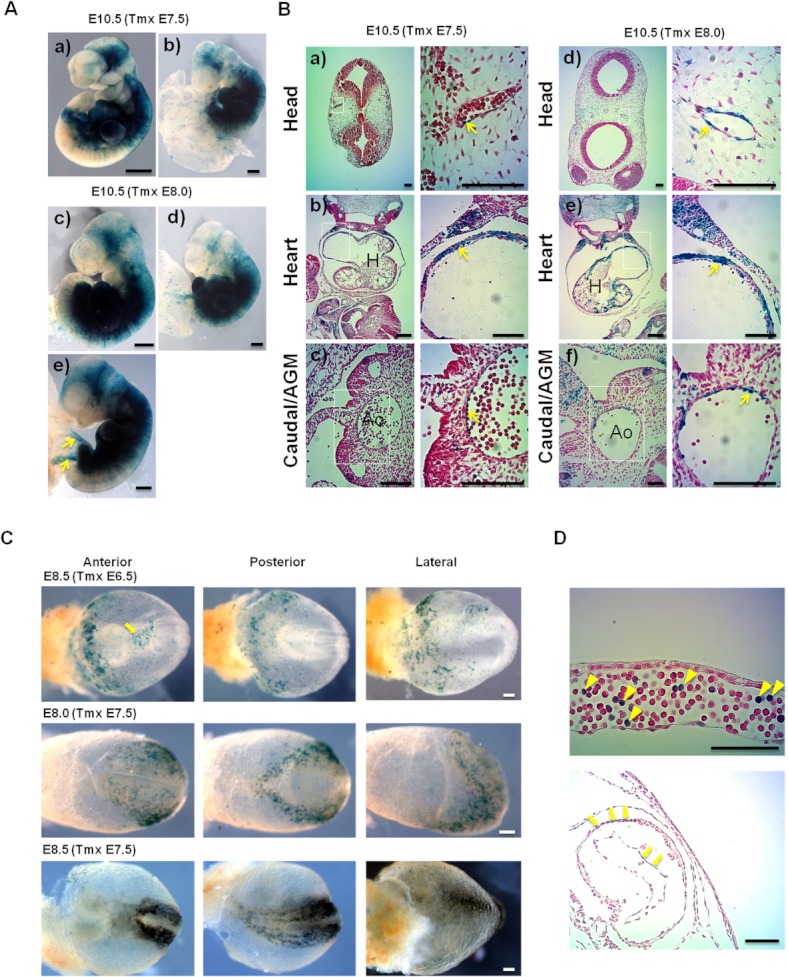
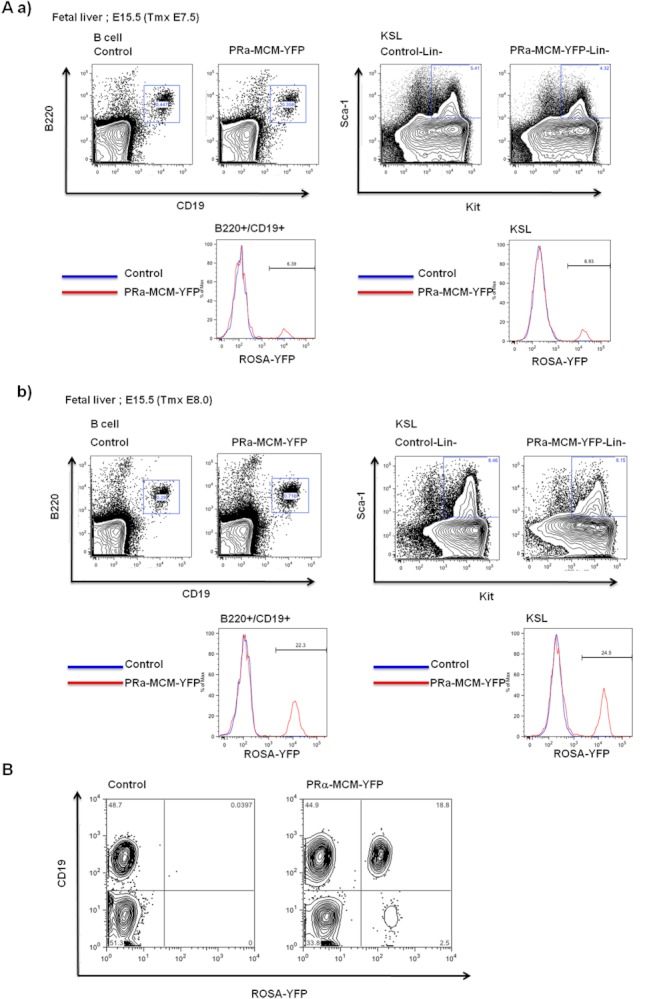
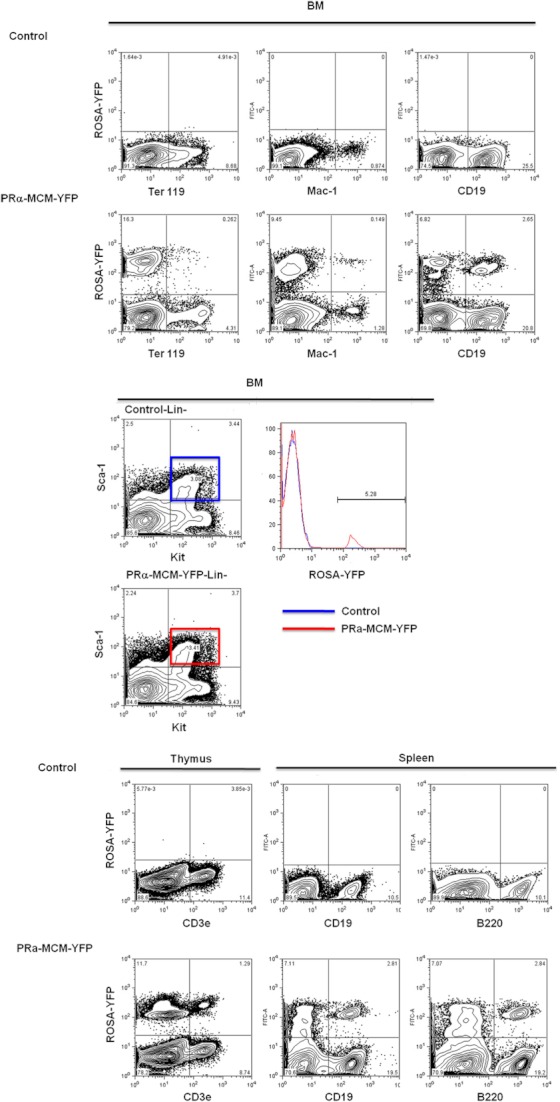
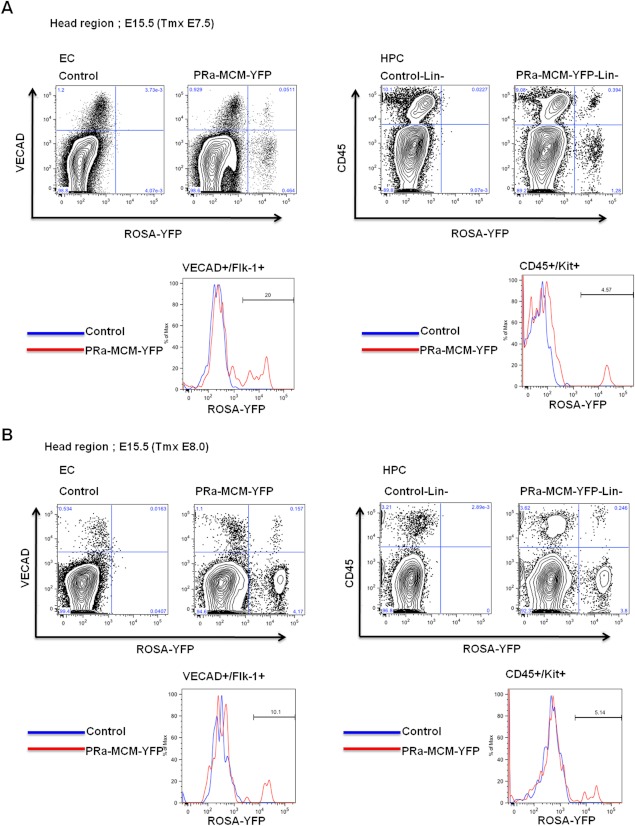
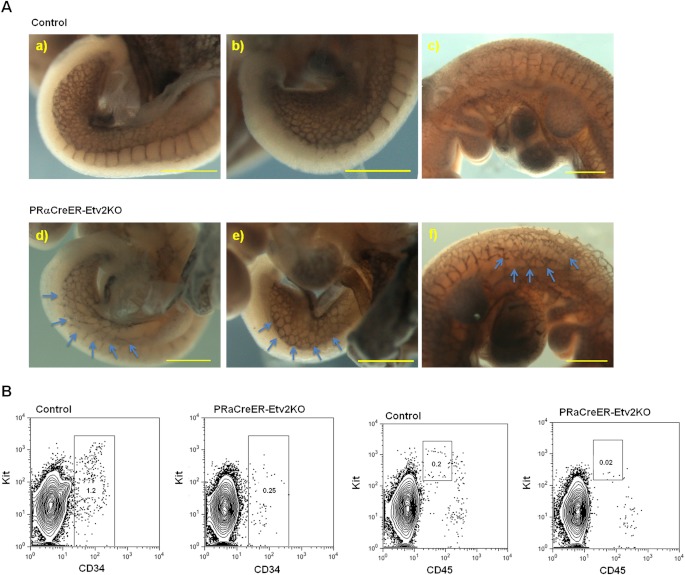
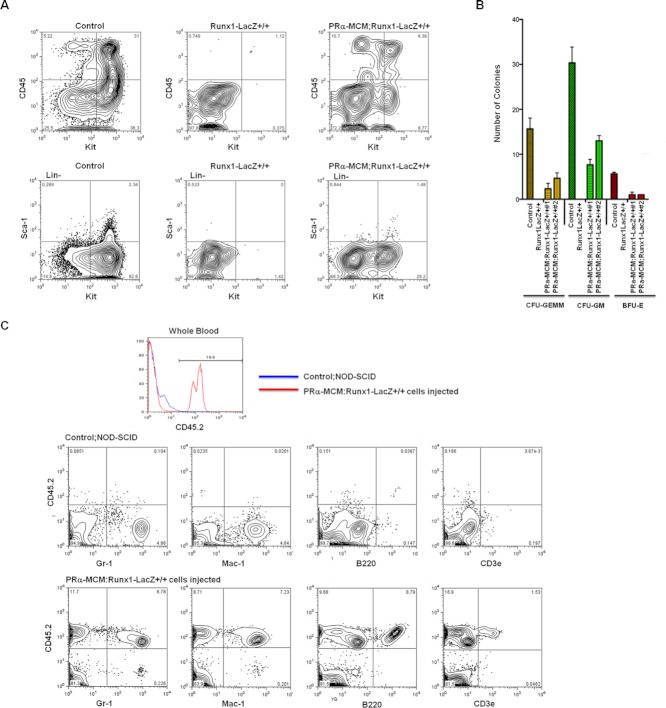
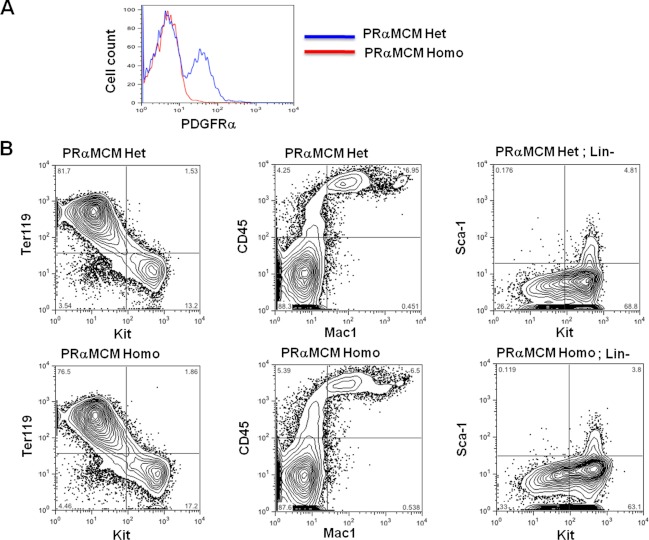
Results: Using PDGFRαMerCreMer mice, PDGFRα+ early mesoderm was shown to contribute to endothelial cells including hemogenic ECs, fetal liver B lymphocytes, and Lin-Kit+Sca-1+ (KSL) cells. Contribution of PDGFRα+ mesoderm into ECs and HPCs was limited until E8.5, indicating that PDGFRα+/Flk-1+ population that exists until E8.5 may be the source for hemato-endothelial lineages from PDGFRα+ population. The functional significance of PDGFRα+ mesoderm in vascular development and hematopoiesis was confirmed by genetic deletion of Etv2 or restoration of Runx1 in PDGFRα+ cells. Etv2 deletion and Runx1 restoration in PDGFRα+ cells resulted in abnormal vascular remodeling and rescue of fetal liver CD45+ and Lin-Kit+Sca-1+ (KSL) cells, respectively.
Conclusions: Endothelial and hematopoietic cells can be derived from PDGFRα+ early mesoderm in mice. PDGFRα+ mesoderm is functionally significant in vascular development and hematopoiesis from phenotype analysis of genetically modified embryos.
Copyright © 2013 Wiley Periodicals, Inc.
Figures

References
-
- Corbel C, Salaun J, Belo-Diabangouaya P, Dieterlen-Lievre F. Hematopoietic potential of the pre-fusion allantois. Dev Biol. 2007;301:478–488. - PubMed
-
- Cumano A, Dieterlen-Lievre F, Godin I. Lymphoid potential, probed before circulation in mouse, is restricted to caudal intraembryonic splanchnopleura. Cell. 1996;86:907–916. - PubMed
-
- Davis RP, Ng ES, Costa M, Mossman AK, Sourris K, Elefanty AG, Stanley EG. Targeting a GFP reporter gene to the MIXL1 locus of human embryonic stem cells identifies human primitive streak-like cells and enables isolation of primitive hematopoietic precursors. Blood. 2008;111:1876–1884. - PubMed
-
- Drake CJ, Fleming PA. Vasculogenesis in the day 6.5 to 9.5 mouse embryo. Blood. 2000;95:1671–1679. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
