

Integration of canonical and noncanonical Wnt signaling pathways patterns the neuroectoderm along the anterior-posterior axis of sea urchin embryos
- PMID: 23335859
- PMCID: PMC3545869
- DOI: 10.1371/journal.pbio.1001467
Integration of canonical and noncanonical Wnt signaling pathways patterns the neuroectoderm along the anterior-posterior axis of sea urchin embryos
Abstract
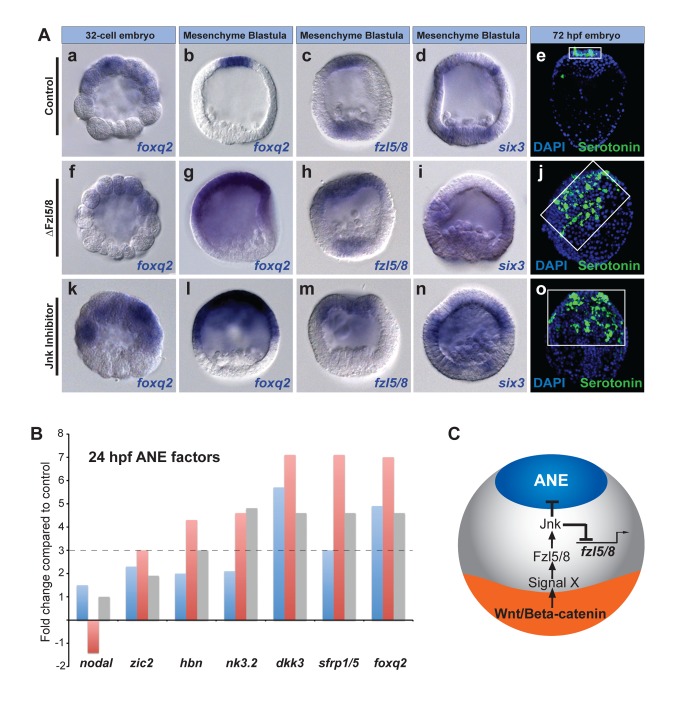
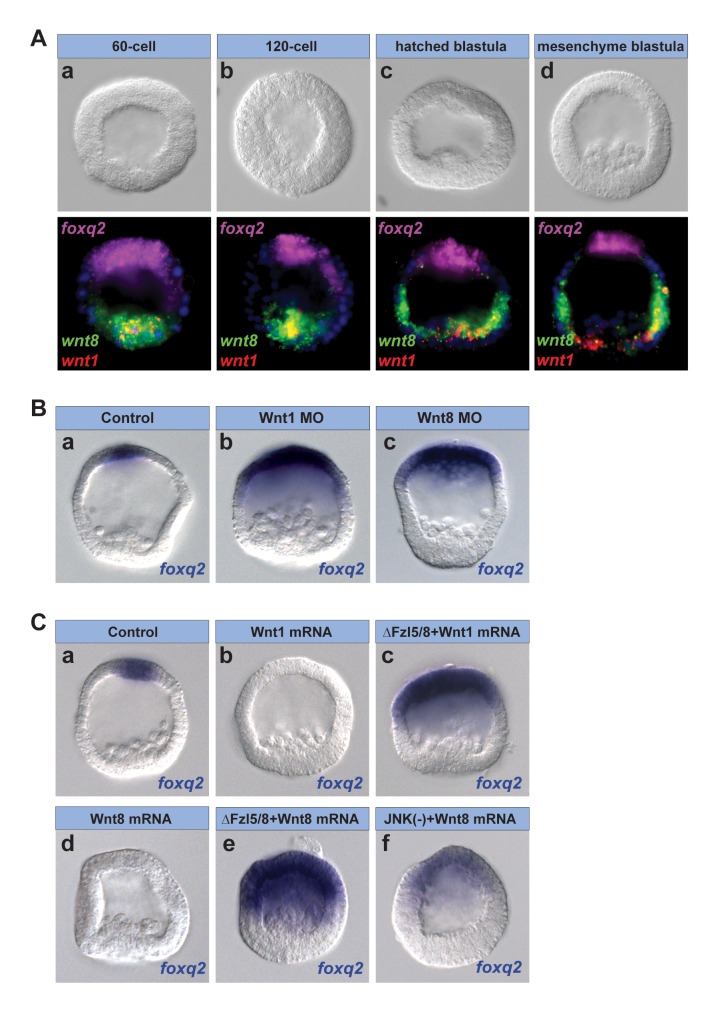
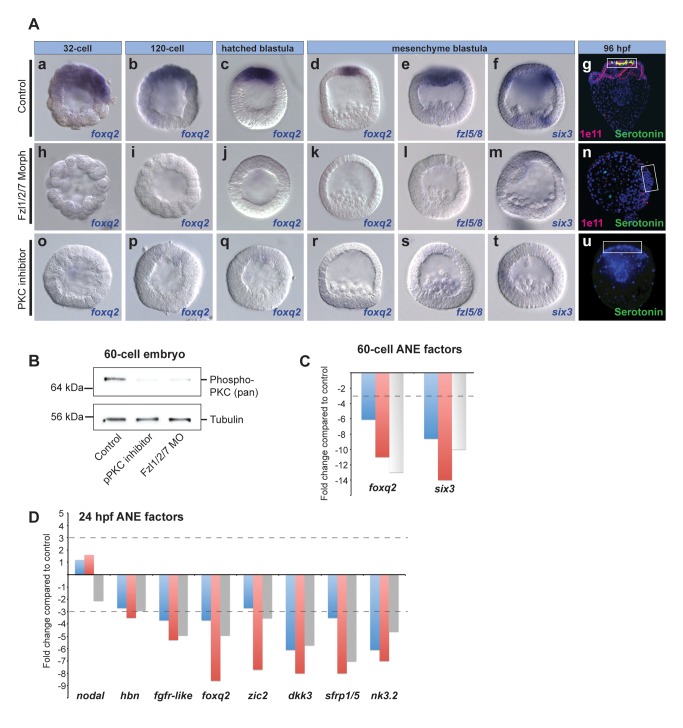
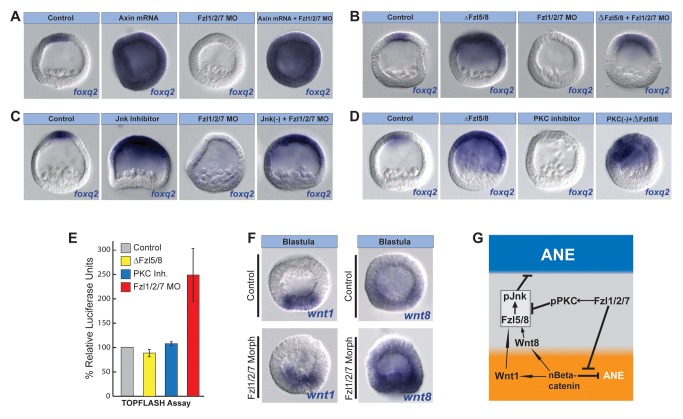
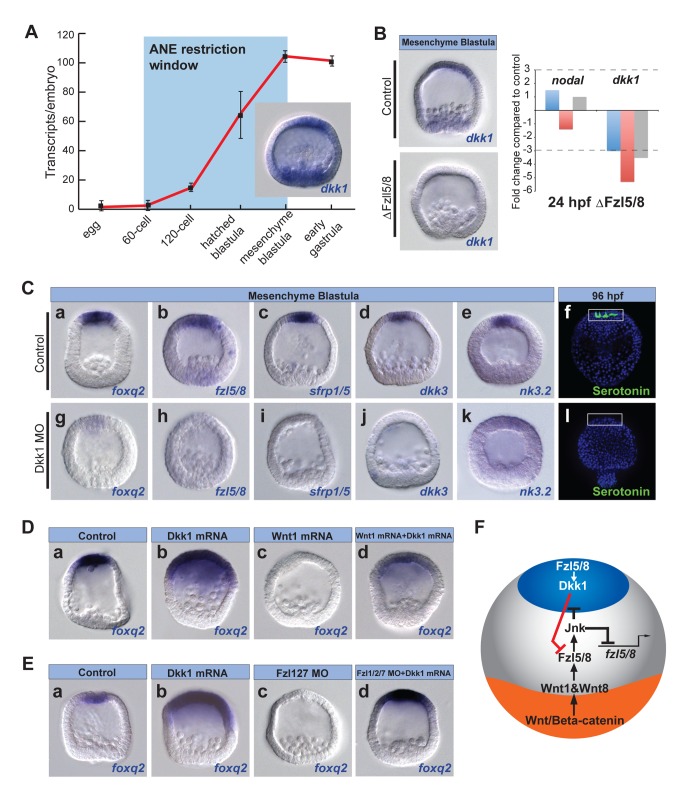
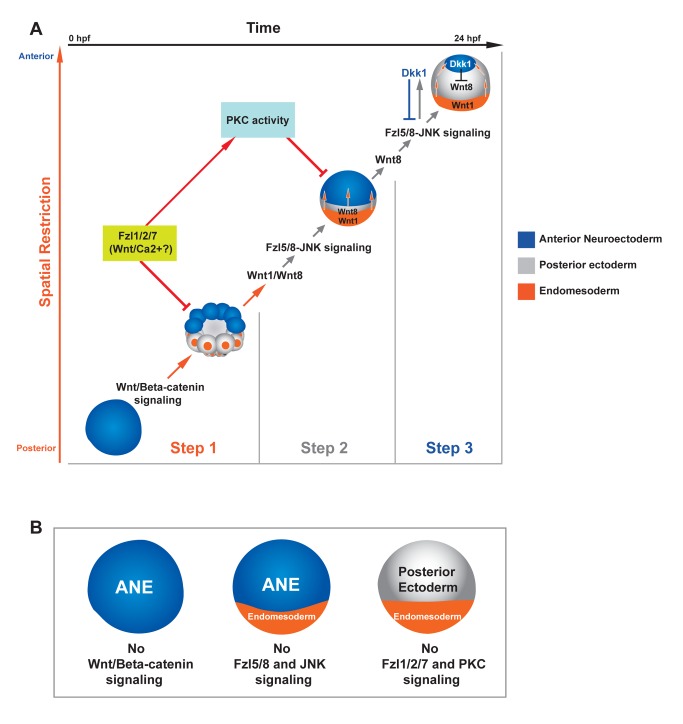
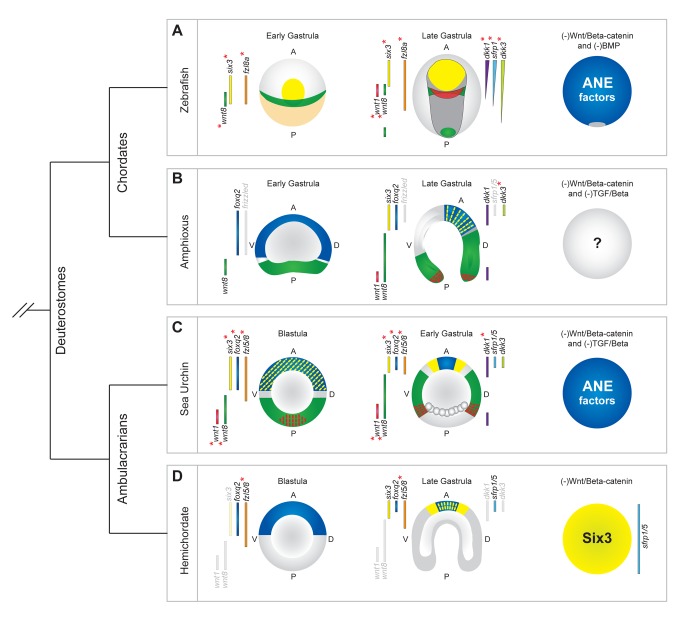
Patterning the neuroectoderm along the anterior-posterior (AP) axis is a critical event in the early development of deuterostome embryos. However, the mechanisms that regulate the specification and patterning of the neuroectoderm are incompletely understood. Remarkably, the anterior neuroectoderm (ANE) of the deuterostome sea urchin embryo expresses many of the same transcription factors and secreted modulators of Wnt signaling, as does the early vertebrate ANE (forebrain/eye field). Moreover, as is the case in vertebrate embryos, confining the ANE to the anterior end of the embryo requires a Wnt/β-catenin-dependent signaling mechanism. Here we use morpholino- or dominant negative-mediated interference to demonstrate that the early sea urchin embryo integrates information not only from Wnt/β-catenin but also from Wnt/Fzl5/8-JNK and Fzl1/2/7-PKC pathways to provide precise spatiotemporal control of neuroectoderm patterning along its AP axis. Together, through the Wnt1 and Wnt8 ligands, they orchestrate a progressive posterior-to-anterior wave of re-specification that restricts the initial, ubiquitous, maternally specified, ANE regulatory state to the most anterior blastomeres. There, the Wnt receptor antagonist, Dkk1, protects this state through a negative feedback mechanism. Because these different Wnt pathways converge on the same cell fate specification process, our data suggest they may function as integrated components of an interactive Wnt signaling network. Our findings provide strong support for the idea that the sea urchin ANE regulatory state and the mechanisms that position and define its borders represent an ancient regulatory patterning system that was present in the common echinoderm/vertebrate ancestor.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Adamska M, Degnan SM, Green KM, Adamski M, Craigie A, et al. (2007) Wnt and TGF-beta expression in the sponge Amphimedon queenslandica and the origin of metazoan embryonic patterning. PloS One 2: e1031 doi:10.1371/journal.pone.0001031. - DOI - PMC - PubMed
-
- Momose T, Derelle R, Houliston E (2008) A maternally localised Wnt ligand required for axial patterning in the cnidarian Clytia hemisphaerica. Development 135: 2105–2113. - PubMed
-
- Logan CY, Miller JR, Ferkowicz MJ, McClay DR (1999) Nuclear beta-catenin is required to specify vegetal cell fates in the sea urchin embryo. Development 126: 345–357. - PubMed
-
- Wikramanayake AH, Hong M, Lee PN, Pang K, Byrum CA, et al. (2003) An ancient role for nuclear beta-catenin in the evolution of axial polarity and germ layer segregation. Nature 426: 446–450. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
