

Stimulus-specific adaptation and deviance detection in the inferior colliculus
- PMID: 23335883
- PMCID: PMC3547232
- DOI: 10.3389/fncir.2012.00089
Stimulus-specific adaptation and deviance detection in the inferior colliculus
Abstract
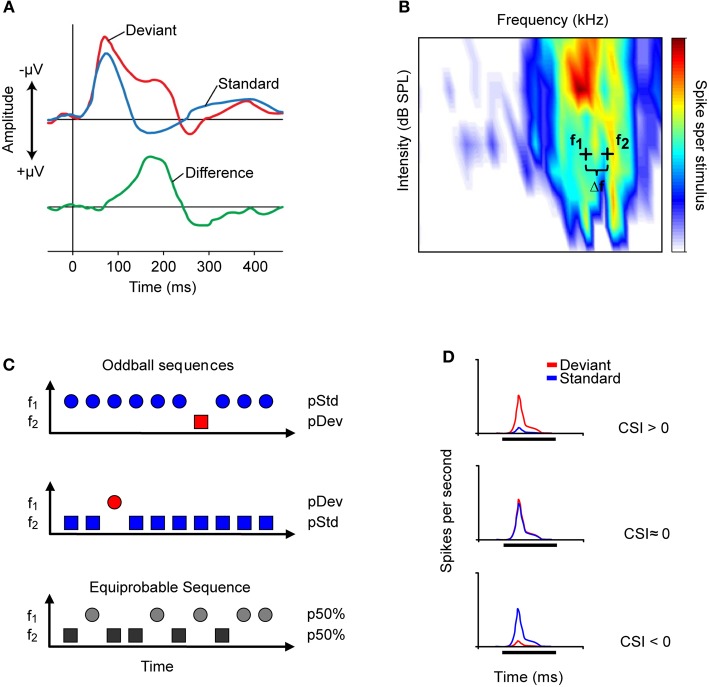
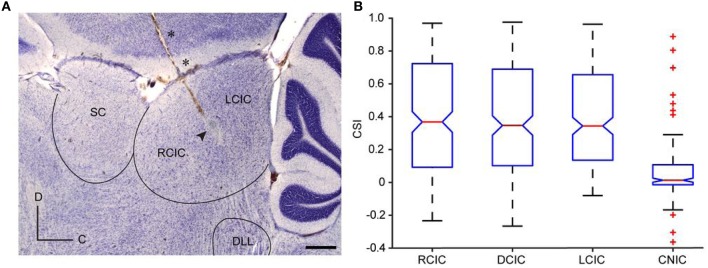
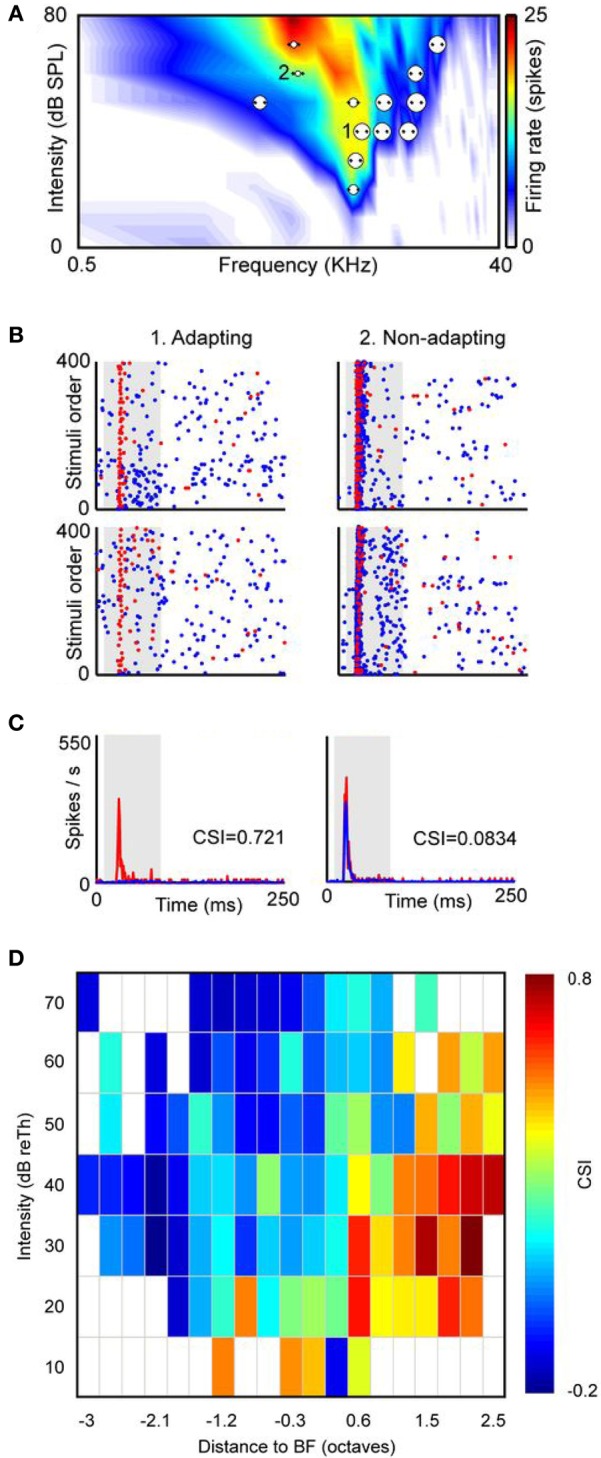
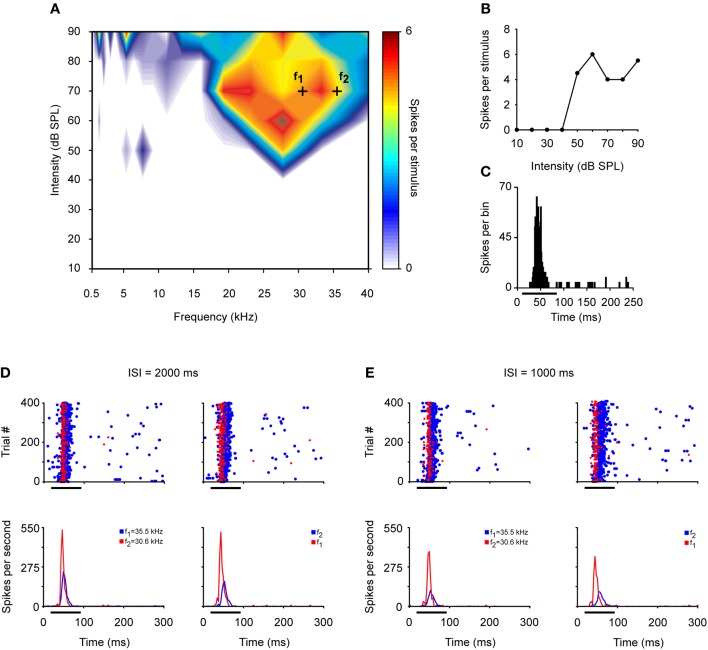
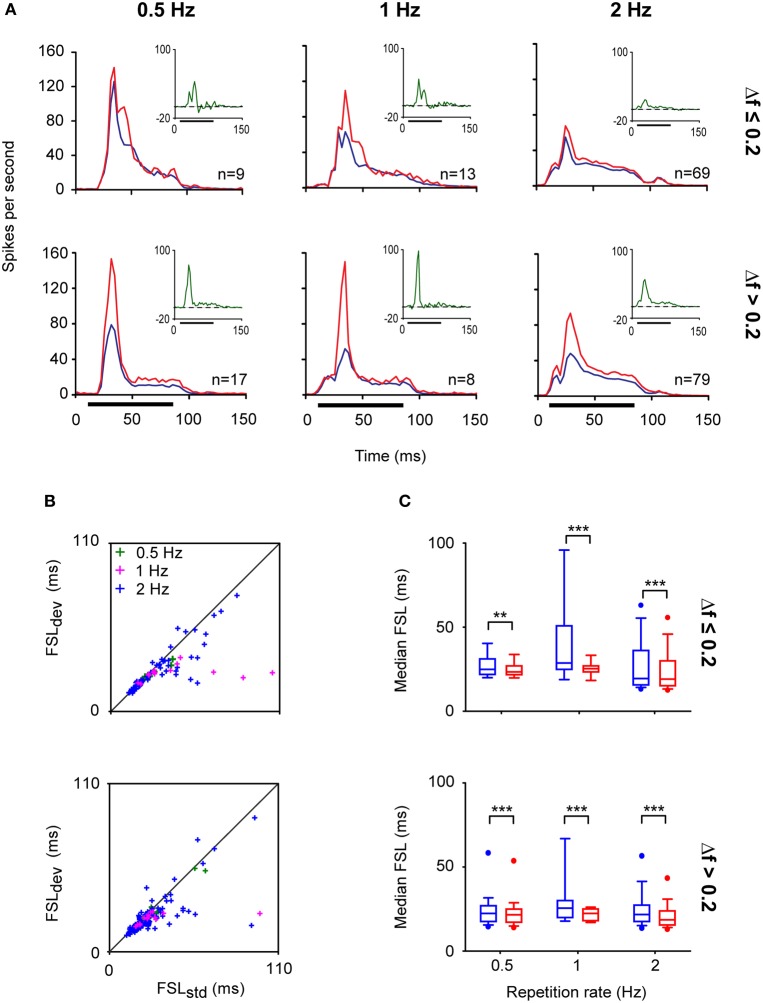
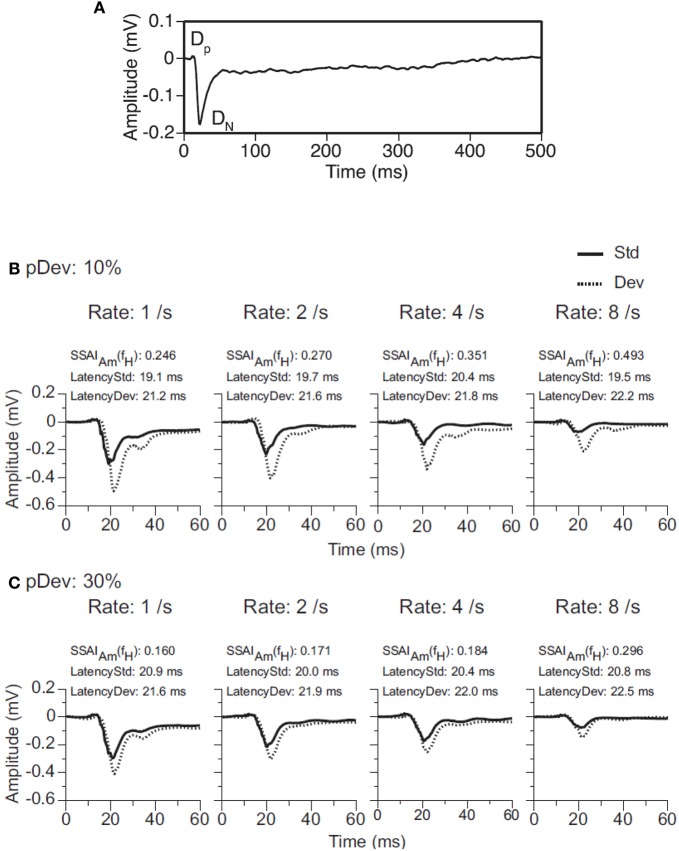
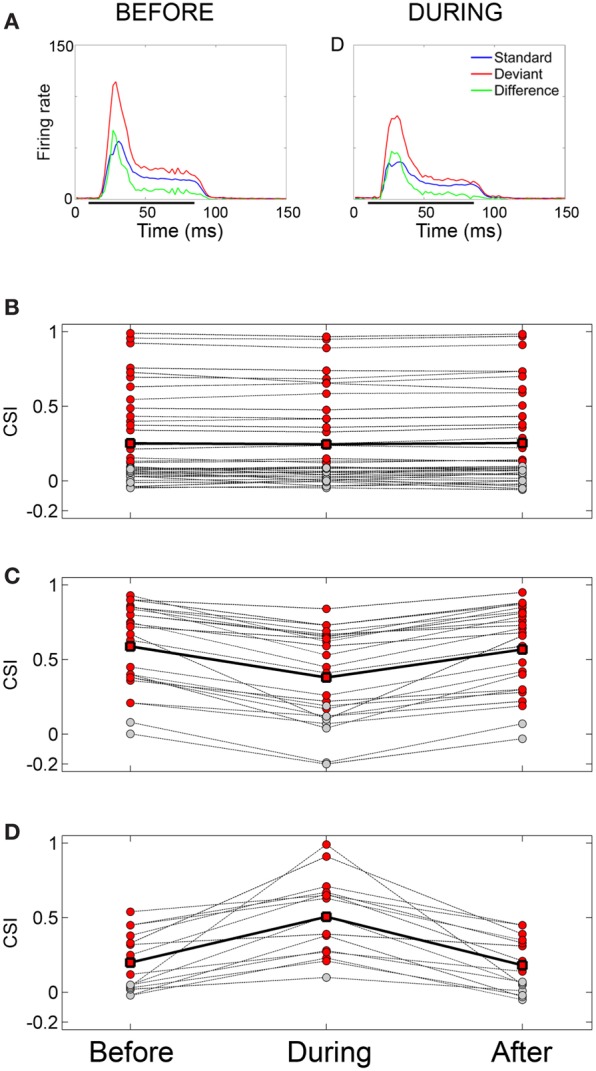
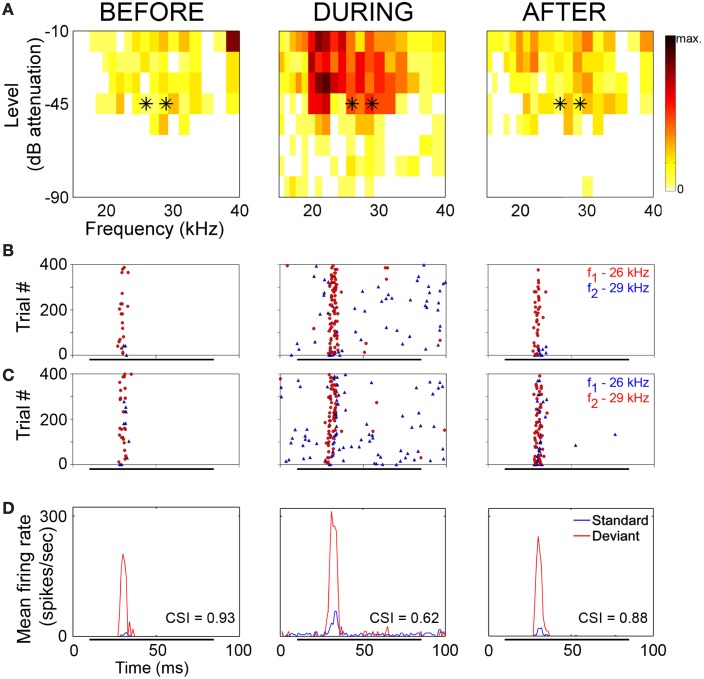
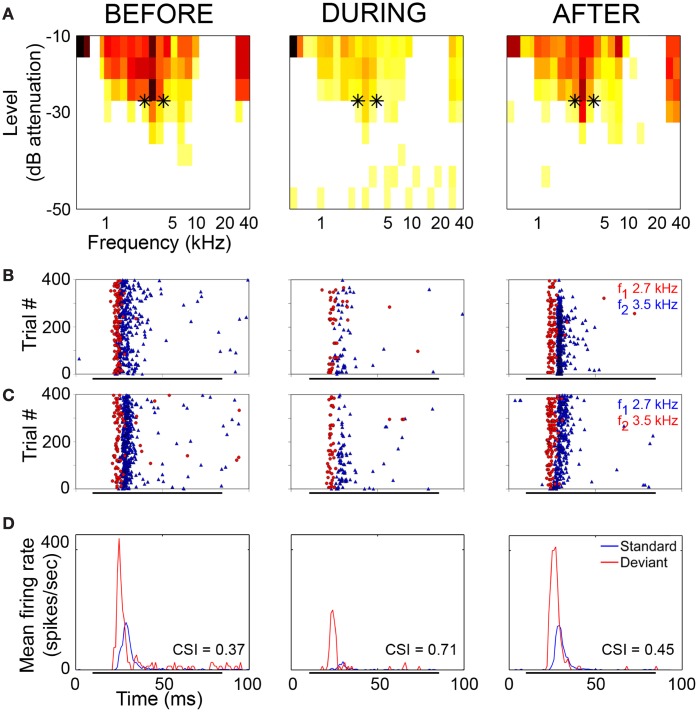
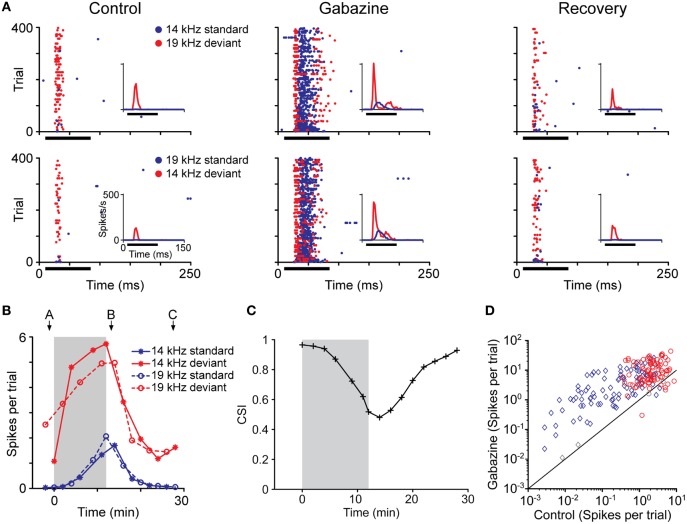
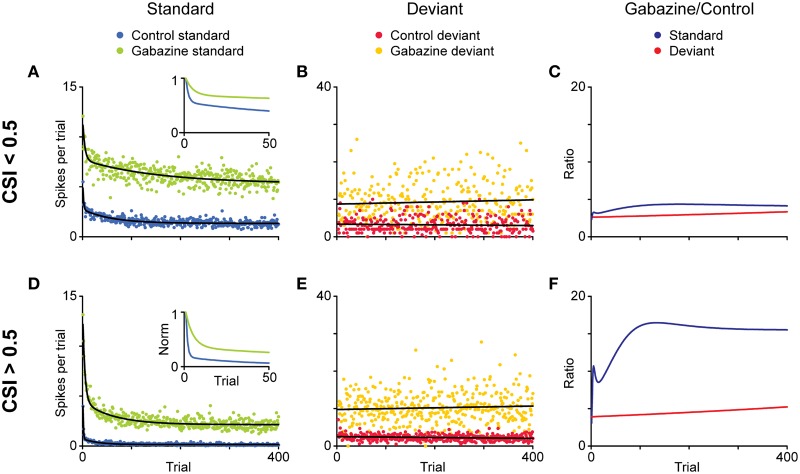
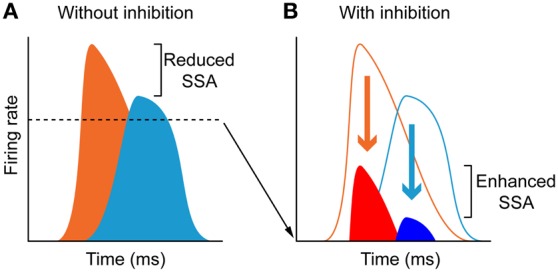
Deviancy detection in the continuous flow of sensory information into the central nervous system is of vital importance for animals. The task requires neuronal mechanisms that allow for an efficient representation of the environment by removing statistically redundant signals. Recently, the neuronal principles of auditory deviance detection have been approached by studying the phenomenon of stimulus-specific adaptation (SSA). SSA is a reduction in the responsiveness of a neuron to a common or repetitive sound while the neuron remains highly sensitive to rare sounds (Ulanovsky et al., 2003). This phenomenon could enhance the saliency of unexpected, deviant stimuli against a background of repetitive signals. SSA shares many similarities with the evoked potential known as the "mismatch negativity," (MMN) and it has been linked to cognitive process such as auditory memory and scene analysis (Winkler et al., 2009) as well as to behavioral habituation (Netser et al., 2011). Neurons exhibiting SSA can be found at several levels of the auditory pathway, from the inferior colliculus (IC) up to the auditory cortex (AC). In this review, we offer an account of the state-of-the art of SSA studies in the IC with the aim of contributing to the growing interest in the single-neuron electrophysiology of auditory deviance detection. The dependence of neuronal SSA on various stimulus features, e.g., probability of the deviant stimulus and repetition rate, and the roles of the AC and inhibition in shaping SSA at the level of the IC are addressed.
Keywords: GABA-mediated inhibition; auditory; change detection; corticofugal modulation; frequency deviance; mismatch negativity; non-lemniscal pathway.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials