

Frequency discrimination and stimulus deviance in the inferior colliculus and cochlear nucleus
- PMID: 23335885
- PMCID: PMC3544151
- DOI: 10.3389/fncir.2012.00119
Frequency discrimination and stimulus deviance in the inferior colliculus and cochlear nucleus
Abstract
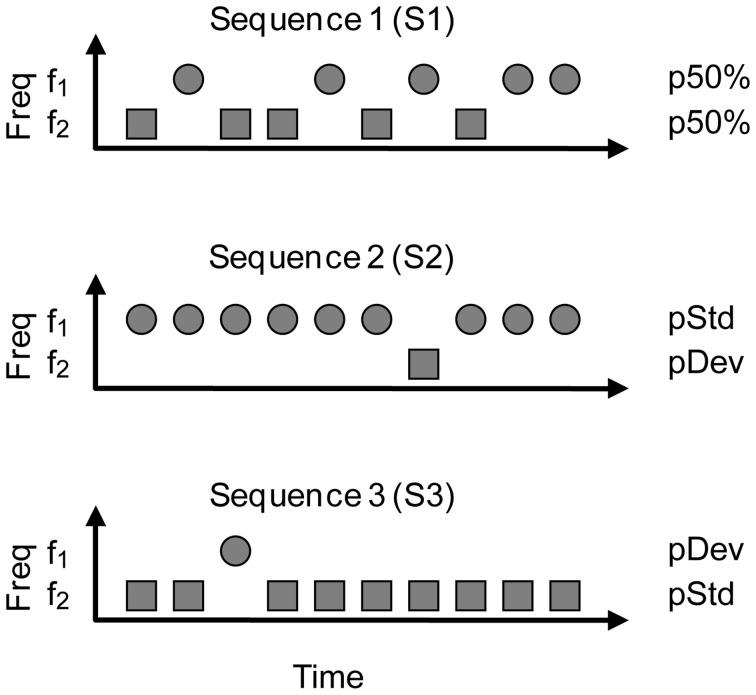
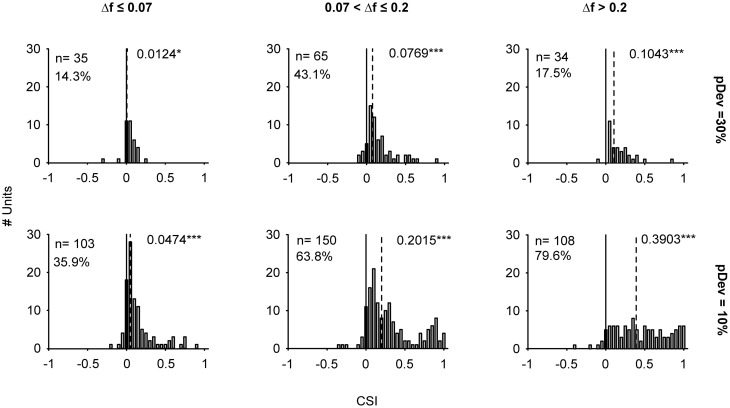
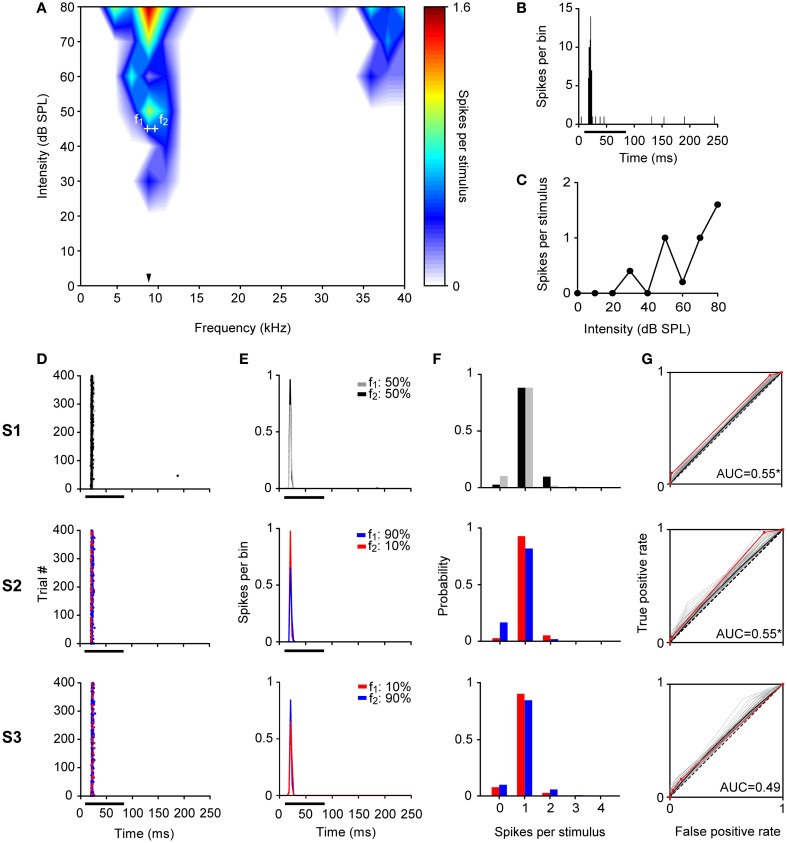
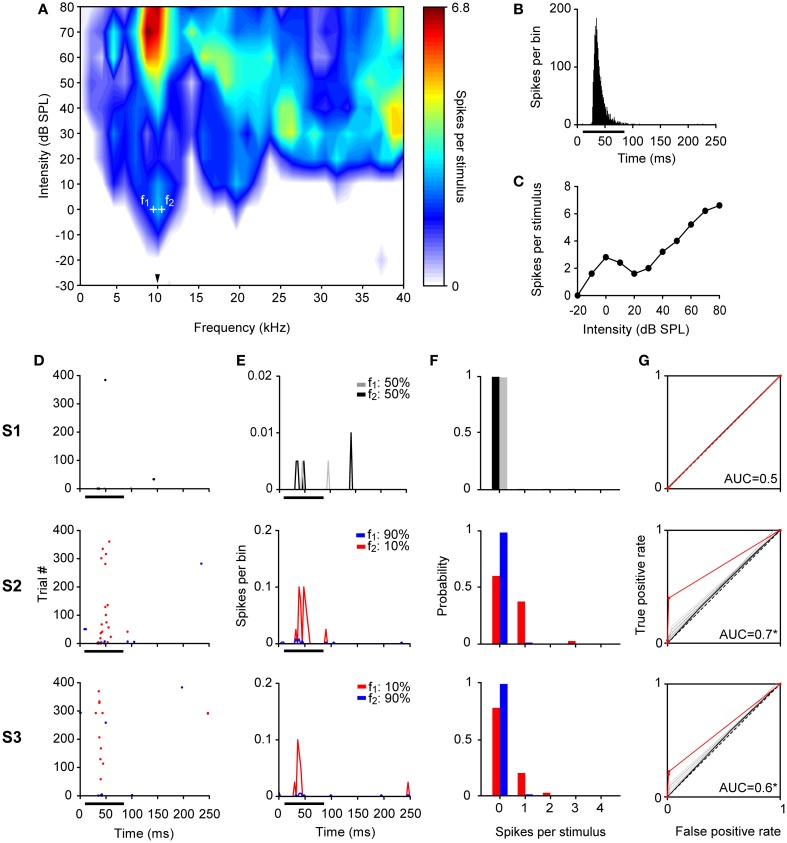
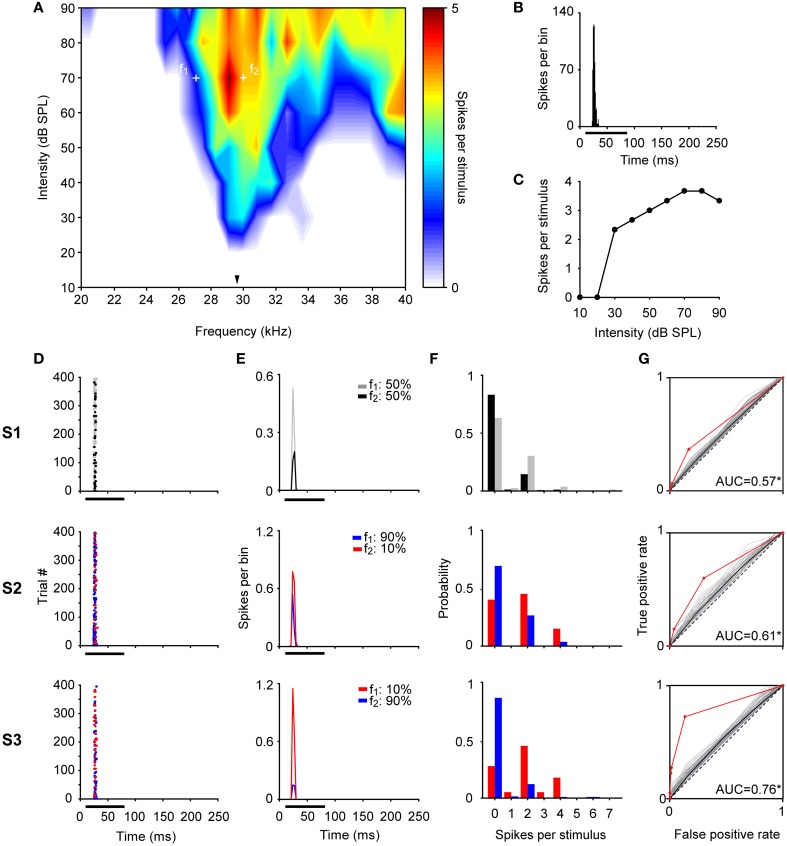
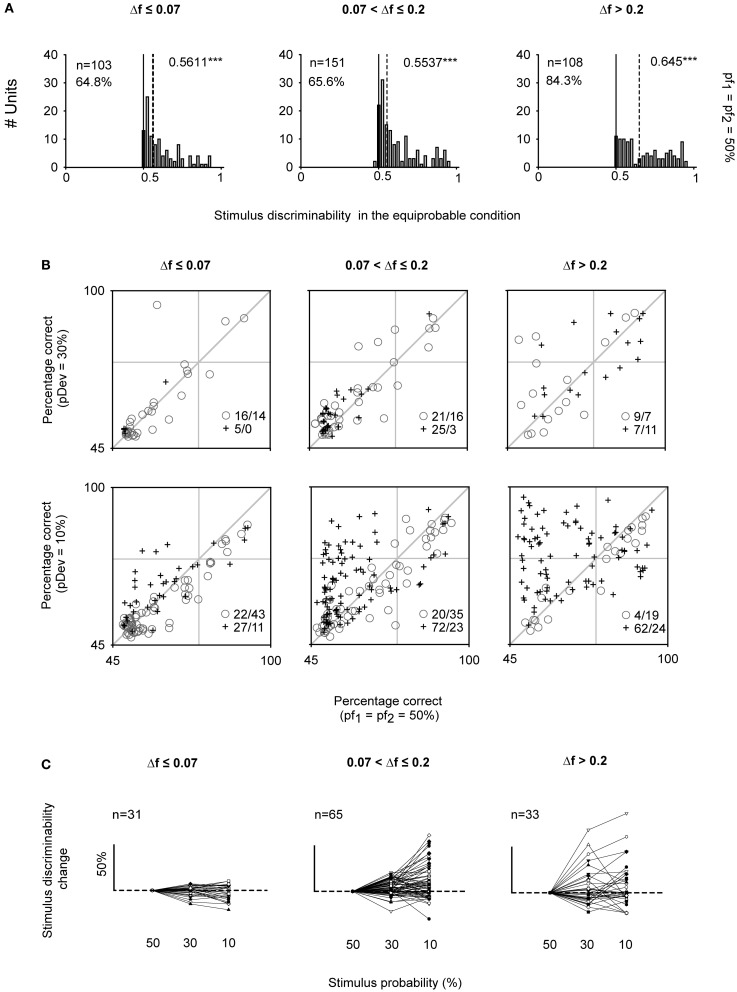
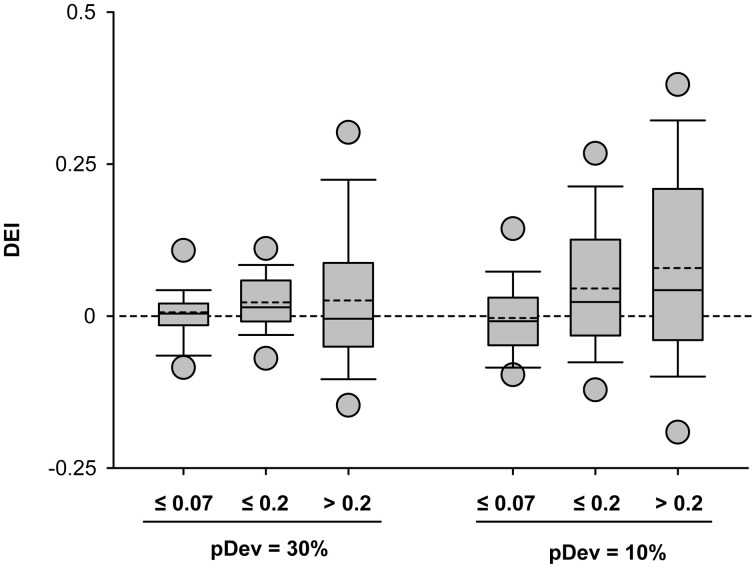
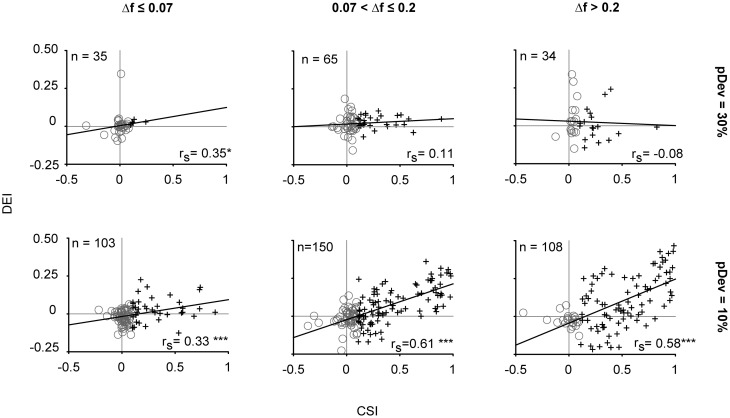
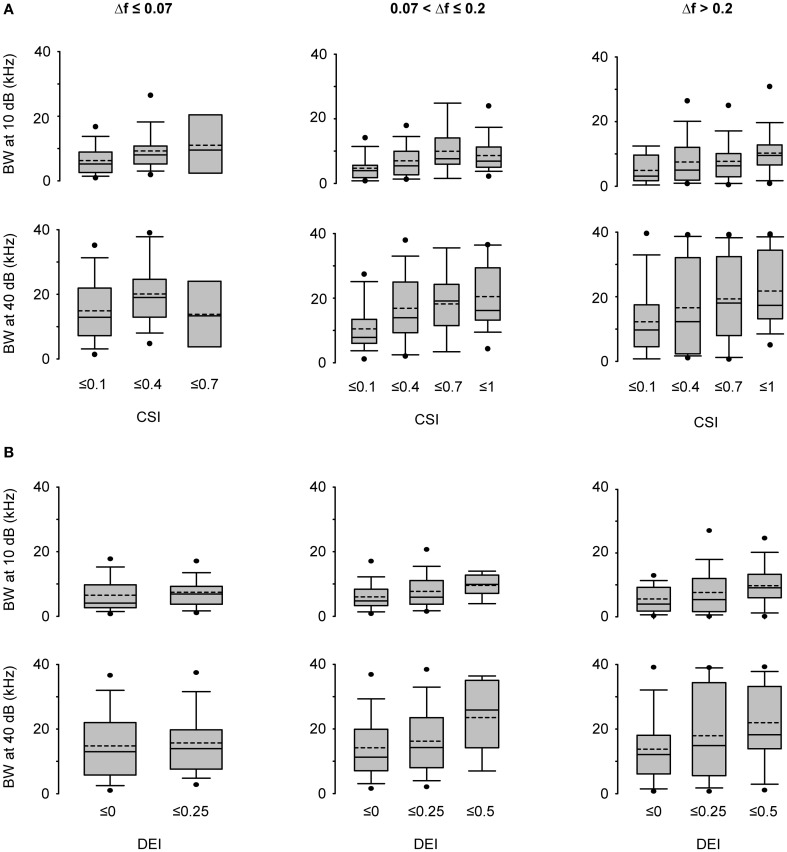
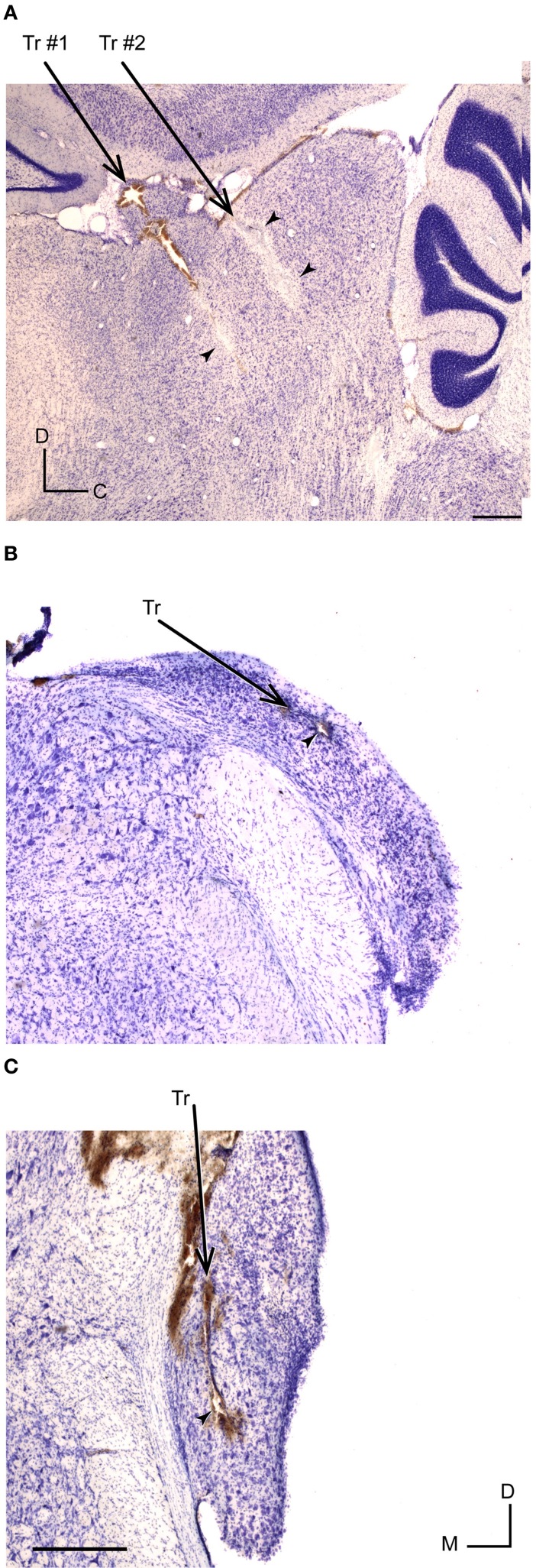
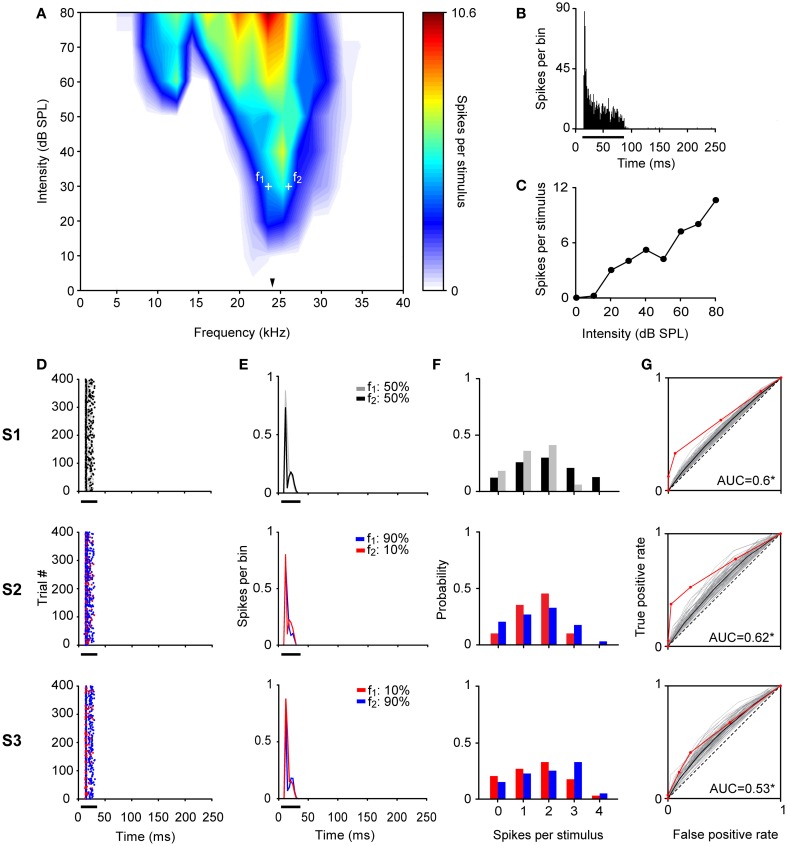
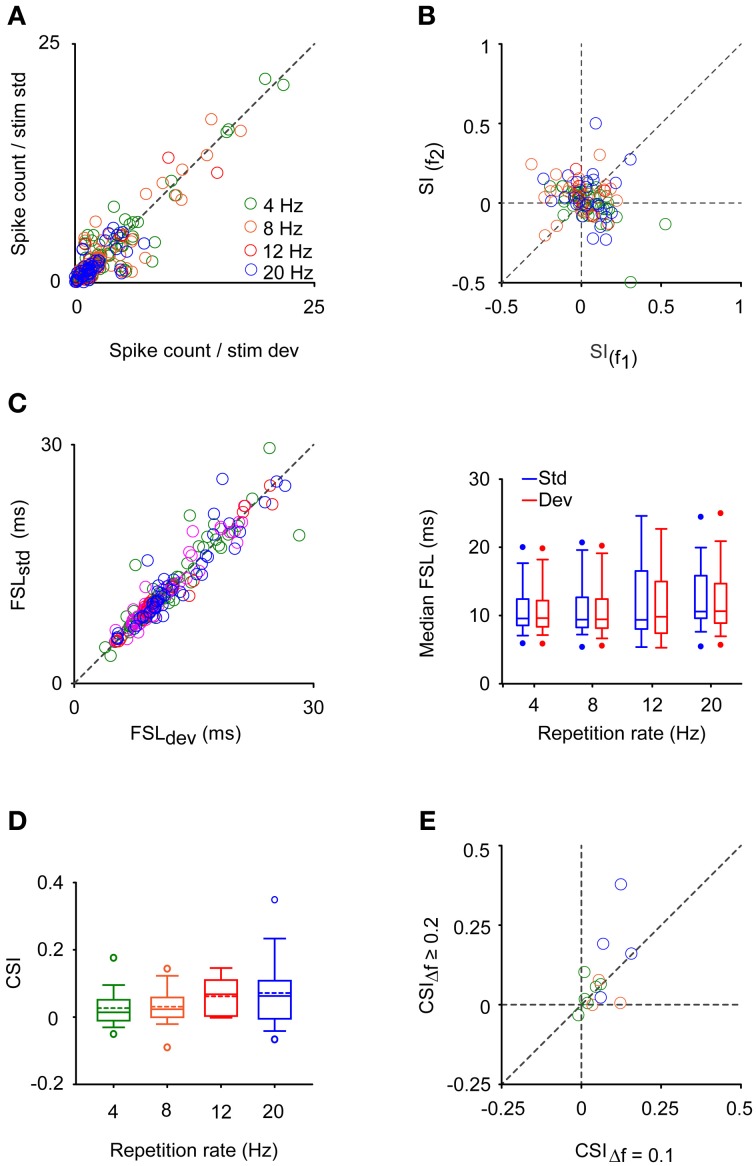
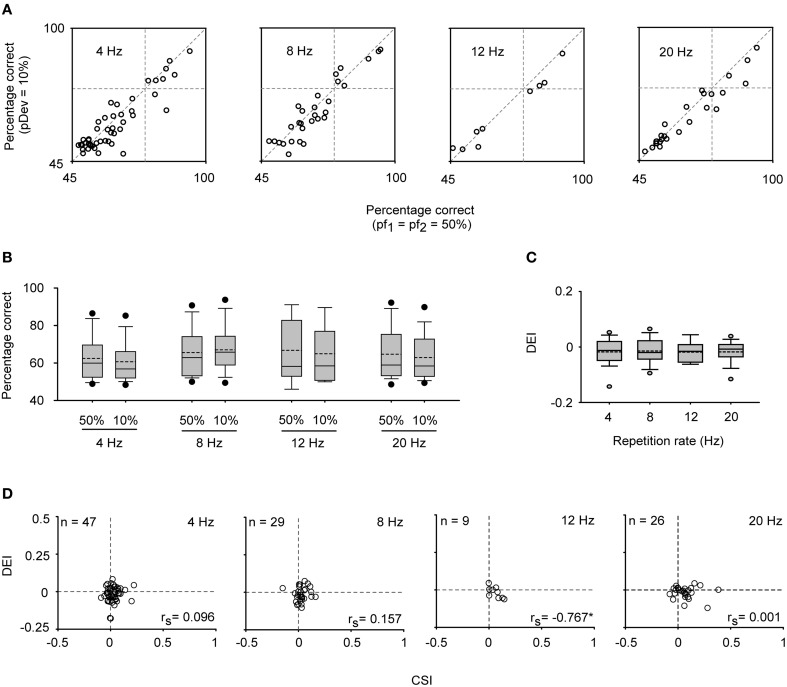
Auditory neurons that exhibit stimulus-specific adaptation (SSA) decrease their response to common tones while retaining responsiveness to rare ones. We recorded single-unit responses from the inferior colliculus (IC) where SSA is known to occur and we explored for the first time SSA in the cochlear nucleus (CN) of rats. We assessed an important functional outcome of SSA, the extent to which frequency discriminability depends on sensory context. For this purpose, pure tones were presented in an oddball sequence as standard (high probability of occurrence) or deviant (low probability of occurrence) stimuli. To study frequency discriminability under different probability contexts, we varied the probability of occurrence and the frequency separation between tones. The neuronal sensitivity was estimated in terms of spike-count probability using signal detection theory. We reproduced the finding that many neurons in the IC exhibited SSA, but we did not observe significant SSA in our CN sample. We concluded that strong SSA is not a ubiquitous phenomenon in the CN. As predicted, frequency discriminability was enhanced in IC when stimuli were presented in an oddball context, and this enhancement was correlated with the degree of SSA shown by the neurons. In contrast, frequency discrimination by CN neurons was independent of stimulus context. Our results demonstrated that SSA is not widespread along the entire auditory pathway, and suggest that SSA increases frequency discriminability of single neurons beyond that expected from their tuning curves.
Keywords: ROC analysis; SSA; change detection; deviant sensitivity; mismatch negativity; non-lemniscal pathway.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials