

Review
doi: 10.1083/jcb.201210007.
Mechanisms of spindle positioning
Affiliations
- PMID: 23337115
- PMCID: PMC3549965
- DOI: 10.1083/jcb.201210007
Item in Clipboard
Review
Mechanisms of spindle positioning
J Cell Biol.
.
Abstract
Accurate positioning of spindles is essential for asymmetric mitotic and meiotic cell divisions that are crucial for animal development and oocyte maturation, respectively. The predominant model for spindle positioning, termed "cortical pulling," involves attachment of the microtubule-based motor cytoplasmic dynein to the cortex, where it exerts a pulling force on microtubules that extend from the spindle poles to the cell cortex, thereby displacing the spindle. Recent studies have addressed important details of the cortical pulling mechanism and have revealed alternative mechanisms that may be used when microtubules do not extend from the spindle to the cortex.
Figures
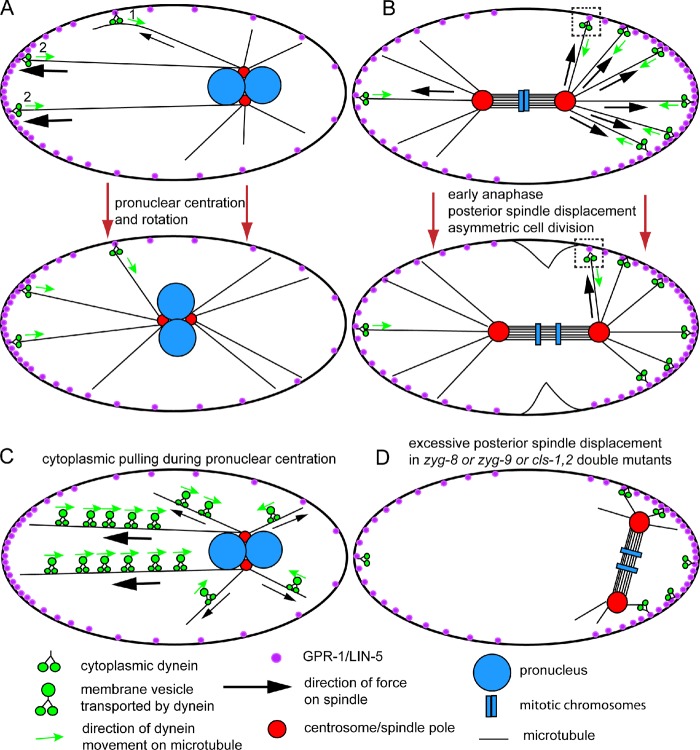
Mitotic spindle movements in the C. elegans zygote. (A) Schematic diagram of a single-celled C. elegans embryo showing cortical pulling by cytoplasmic dynein during pronuclear centration and rotation. The nuclei move toward the anterior (left) so that the spindle assembles in the center of the embryo. (B) Schematic diagram of a single-celled C. elegans embryo showing cortical pulling by dynein during anaphase. The spindle moves to the posterior (right) so that cytokinesis generates two cells of different sizes. The squares highlight a dynein molecule that pulls toward the posterior before spindle displacement, then pulls toward the anterior after spindle displacement. (C) Schematic drawing of cytoplasmic pulling that contributes to centering the pronuclei. (D) Illustration of a spindle that was centered at metaphase but in which both poles moved all the way to the posterior end of the embryo. This occurs in zyg-8 mutants (Gönczy et al., 2001), cls-1,2 (RNAi) embryos (Espiritu et al., 2012), and zyg-9(ts) mutants shifted to a nonpermissive temperature at metaphase (Bellanger et al., 2007), possibly because astral microtubules are too short to reach force generators that would pull toward the anterior.
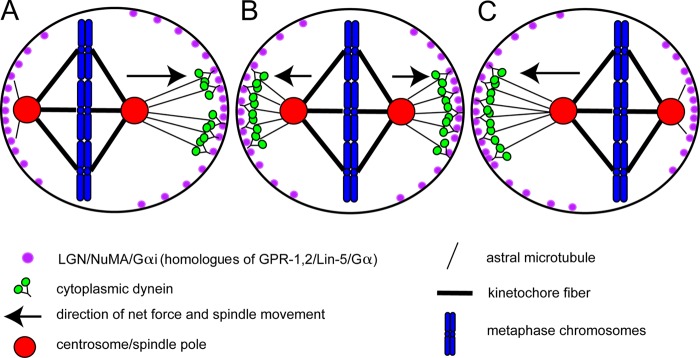
How to center a spindle. Schematic diagram of a metaphase HeLa cell where the spindle oscillates along its pole–pole axis to maintain a centered position to allow symmetric cytokinesis. (A) When the left spindle pole is close to the cortex, Plk1 on the pole (red) causes dynein (green) to dissociate from LGN–NuMA complexes (purple; human homologues of GPR-1,2/LIN-5). (B and C) The spindle moves to the right because of the higher concentration of LGN–NuMA–dynein complexes on the right cortex. When chromosomes are close to the cortex as in A, the GTP-Ran gradient from the chromosomes causes dissociation of LGN/NuMA from the cortex. This second system centers the spindle in the axis perpendicular to the pole–pole axis.
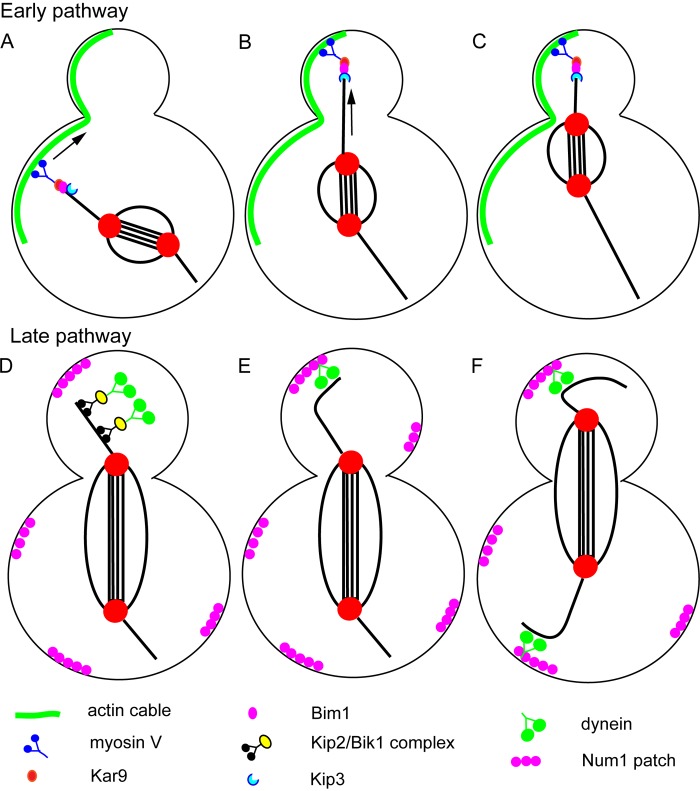
Spindle positioning in budding yeast. Schematic diagram of the two sequential spindle positioning pathways of budding yeast. In the early pathway (A–C), myosin V transports the plus end of an astral microtubule toward the bud tip on a polarized actin cable. Once the plus end has reached the bud cortex, the plus-end depolymerase, KIP3, is activated to allow pulling of the spindle pole toward the bud neck. In the late pathway (D–F), the plus end–directed microtubule motor Kip2 transports dynein to the plus ends of microtubules via the adaptor protein Bik1 (D). Dynein can be targeted to plus ends by two additional Bik1-dependent mechanisms (see text). When dynein reaches the bud cortex on a polymerizing microtubule plus end (E), contact with the cortical protein Num1 allows dynein to pull the spindle toward the bud (F). During early anaphase (D and E), dynein is not loaded onto microtubules in the mother cell. During late anaphase (F), dynein is loaded on microtubules in the mother cell to prevent movement of the spindle all the way into the bud.
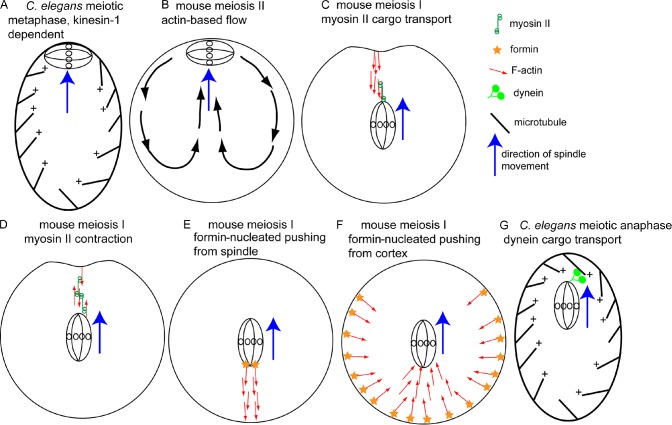
A plethora of nonastral spindle positioning mechanisms. (A) Metaphase C. elegans meiotic spindles are positioned in a parallel orientation at the cortex by microtubules and kinesin-1. (B) The mouse metaphase II spindle may be positioned by actin-dependent cytoplasmic streaming. Pole-first migration of the mouse meiosis I spindle to the cortex may be mediated by cargo transport on parallel actin filaments by spindle pole–bound myosin II (C), myosin II–based contraction of anti-parallel actin filaments (D), or pushing forces generated by polymerizing actin filaments nucleated by formin molecules on the spindle (E) or nucleated by formin molecules on the cortex (F). Red arrows indicate the pointed ends of actin filaments. (G) One spindle pole of the early anaphase C. elegans meiotic spindle may be transported to the cortex as cargo by dynein on polarized cytoplasmic microtubules.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
