

Testosterone alters maternal vascular adaptations: role of the endothelial NO system
- PMID: 23339170
- PMCID: PMC3596870
- DOI: 10.1161/HYPERTENSIONAHA.111.00486
Testosterone alters maternal vascular adaptations: role of the endothelial NO system
Abstract
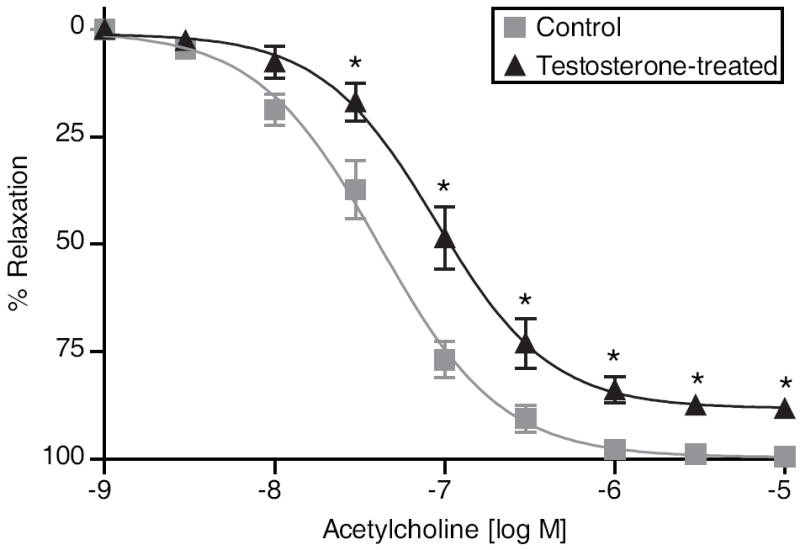
Sex steroid hormones estradiol and progesterone play an important role in vascular adaptations during pregnancy. However, little is known about the role of androgens. Plasma testosterone (T) levels are elevated in preeclampsia, mothers with polycystic ovary, and pregnant African American women, who have endothelial dysfunction and develop gestational hypertension. We tested whether increased T alters vascular adaptations during pregnancy and whether these alterations depend on endothelium-derived factors, such as prostacyclin, endothelium-derived hyperpolarizing factor, and NO. Pregnant Sprague Dawley rats were injected with vehicle (n=12) or T propionate [0.5 mg/Kg per day from gestation day 15-19; n=12] to increase plasma T levels 2-fold, similar to that observed in preeclampsia. Telemetric blood pressures and endothelium-dependent vascular reactivity were assessed with wire-myograph system. Phospho-endothelial NO synthase and total endothelial NO synthase were examined in mesenteric arteries. Mean arterial pressures were significantly higher starting from gestation day19 until delivery in T-treated dams. Endothelium-dependent relaxation responses to acetylcholine were significantly lower in mesenteric arteries of T-treated dams (pD(2) [-log EC(50)]=7.05±0.06; E(max)=89.4±1.89) compared with controls (pD(2)=7.38±0.04; E(max)=99.9±0.97). Further assessment of endothelial factors showed NO-mediated relaxations were blunted in T-treated mesenteric arteries (E(max)=42.26±5.95) compared with controls (E(max)=76.49±5.06); however, prostacyclin- and endothelium-derived hyperpolarizing factor-mediated relaxations were unaffected. Relaxation to sodium nitroprusside was unaffected with T-treatment. Phosphorylations of endothelial NO synthase at Ser(1177) were decreased and at Thr(495) increased in T-treated mesenteric arteries without changes in total endothelial NO synthase levels. In conclusion, increased maternal T, at concentrations relevant to abnormal clinical conditions, cause hypertension associated with blunting of NO-mediated vasodilation. T may induce the increased vascular resistance associated with pregnancy-induced hypertension.
Conflict of interest statement
None
Figures





Comment in
-
Testosterone and sympathetic nerve activity during pregnancy.Hypertension. 2013 May;61(5):e44. doi: 10.1161/HYPERTENSIONAHA.113.01193. Epub 2013 Mar 25. Hypertension. 2013. PMID: 23529174 No abstract available.
-
Response to Testosterone and sympathetic nerve activity during pregnancy.Hypertension. 2013 May;61(5):e45. doi: 10.1161/hypertensionaha.113.01216. Hypertension. 2013. PMID: 23724418 No abstract available.
References
-
- Thornburg KL, Jacobson SL, Giraud GD, Morton MJ. Hemodynamic changes in pregnancy. Semin Perinatol. 2000;24:11–14. - PubMed
-
- Magness RR. Maternal cardiovascular and other physiologic responses to the endocrinology of pregnancy. In: Bazer FW, editor. The Endocrinology of Pregnancy. Totowa, NJ: Humana Press Inc.; 1998. pp. 507–539.
-
- Conrad KP, Joffe GM, Kruszyna H, Kruszyna R, Rochelle LG, Smith RP, Chavez JE, Mosher MD. Identification of increased nitric oxide biosynthesis during pregnancy in rats. FASEB J. 1993;7:566–571. - PubMed
-
- Williams DJ, Vallance PJ, Neild GH, Spencer JA, Imms FJ. Nitric oxide-mediated vasodilation in human pregnancy. Am J Physiol. 1997;272:H748–H752. - PubMed
-
- Sladek SM, Magness RR, Conrad KP. Nitric oxide and pregnancy. Am J Physiol. 1997;272:R441–R463. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
