

Orchestration of thiamin biosynthesis and central metabolism by combined action of the thiamin pyrophosphate riboswitch and the circadian clock in Arabidopsis
- PMID: 23341335
- PMCID: PMC3584542
- DOI: 10.1105/tpc.112.106385
Orchestration of thiamin biosynthesis and central metabolism by combined action of the thiamin pyrophosphate riboswitch and the circadian clock in Arabidopsis
Abstract
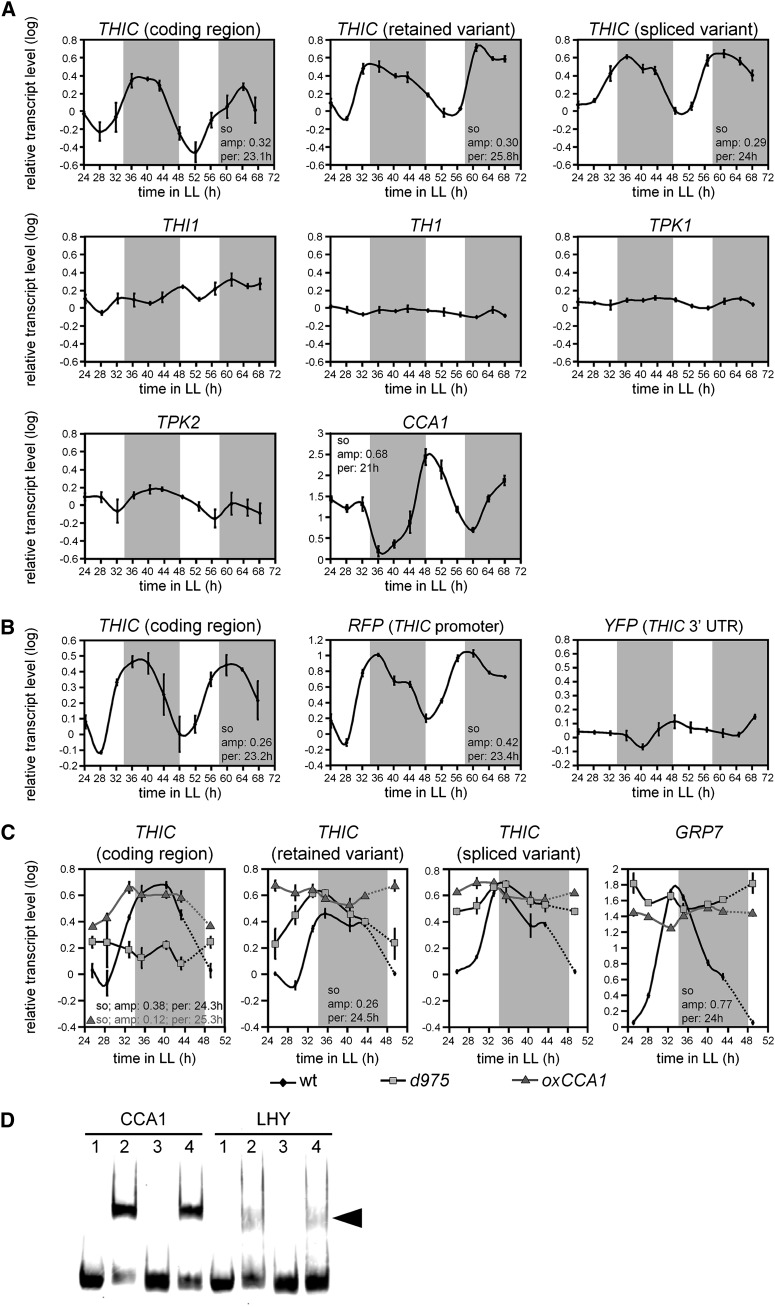
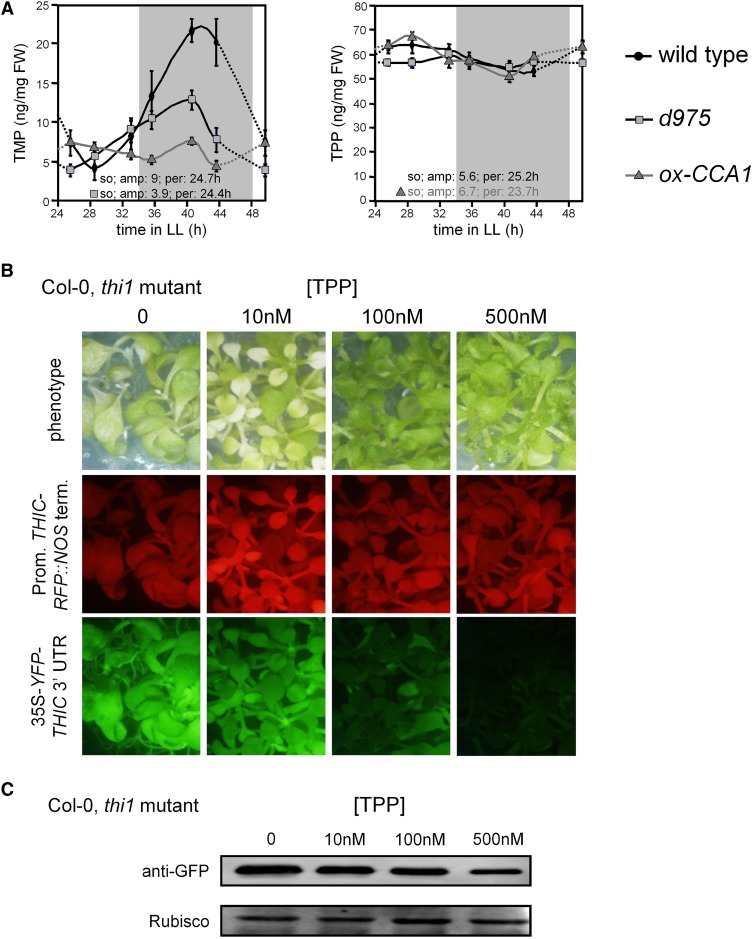
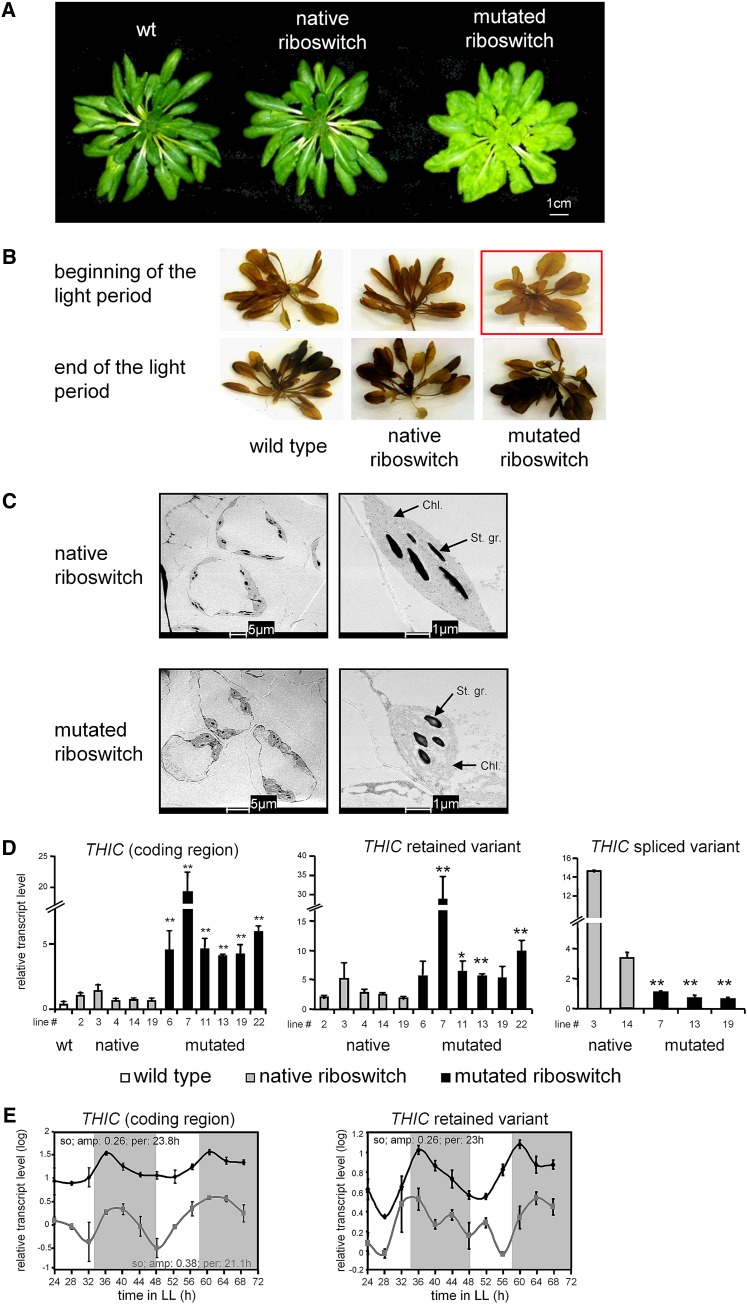
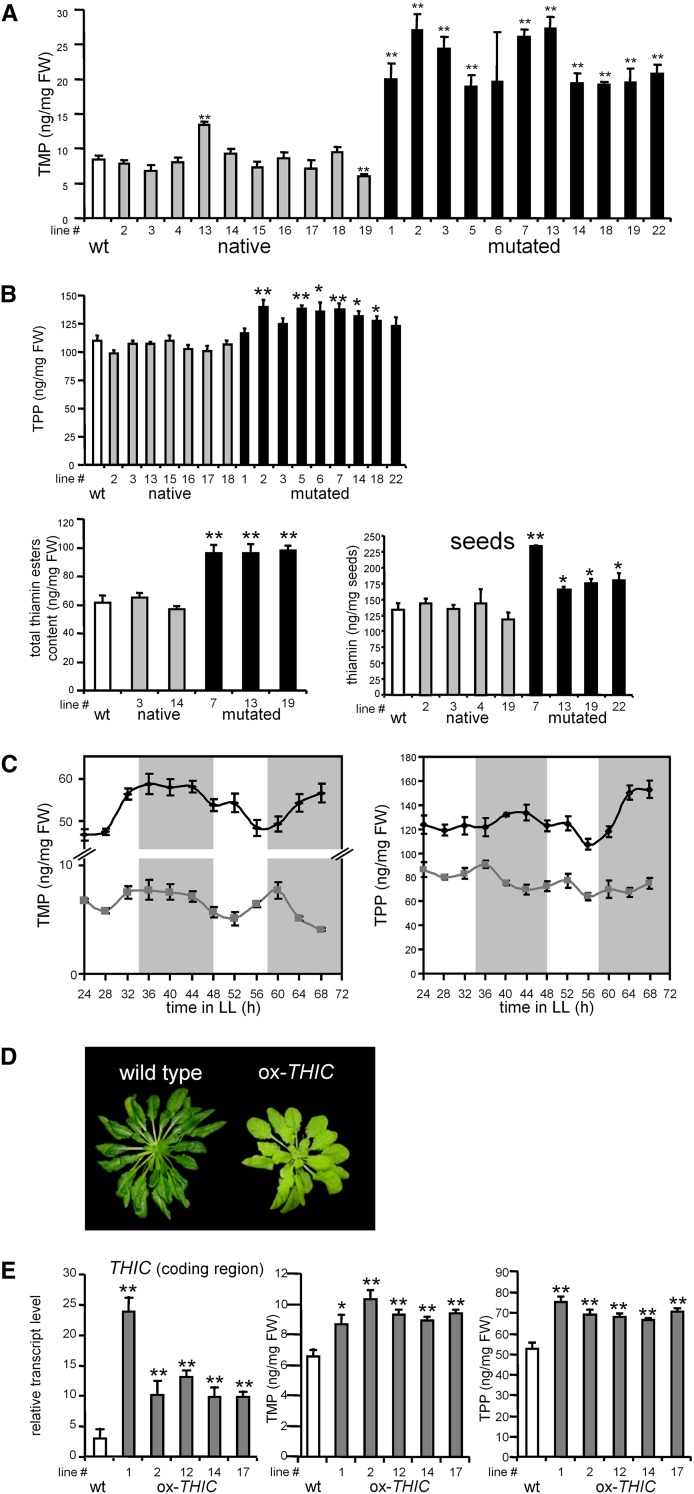
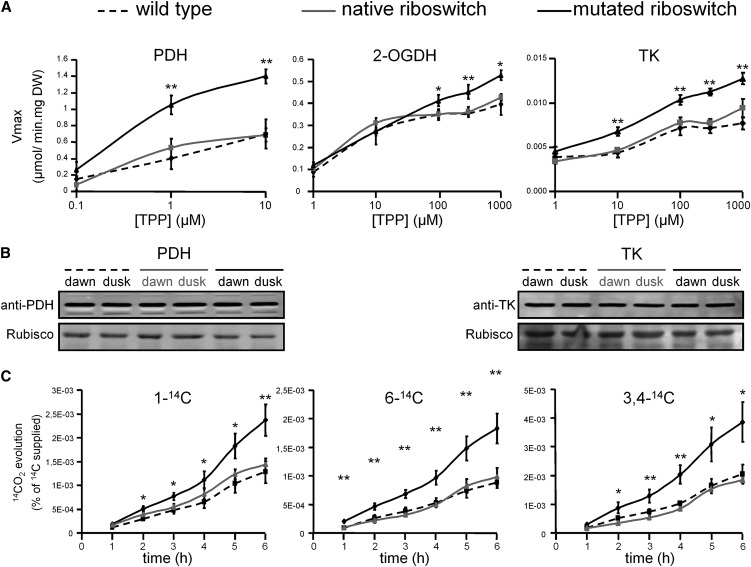
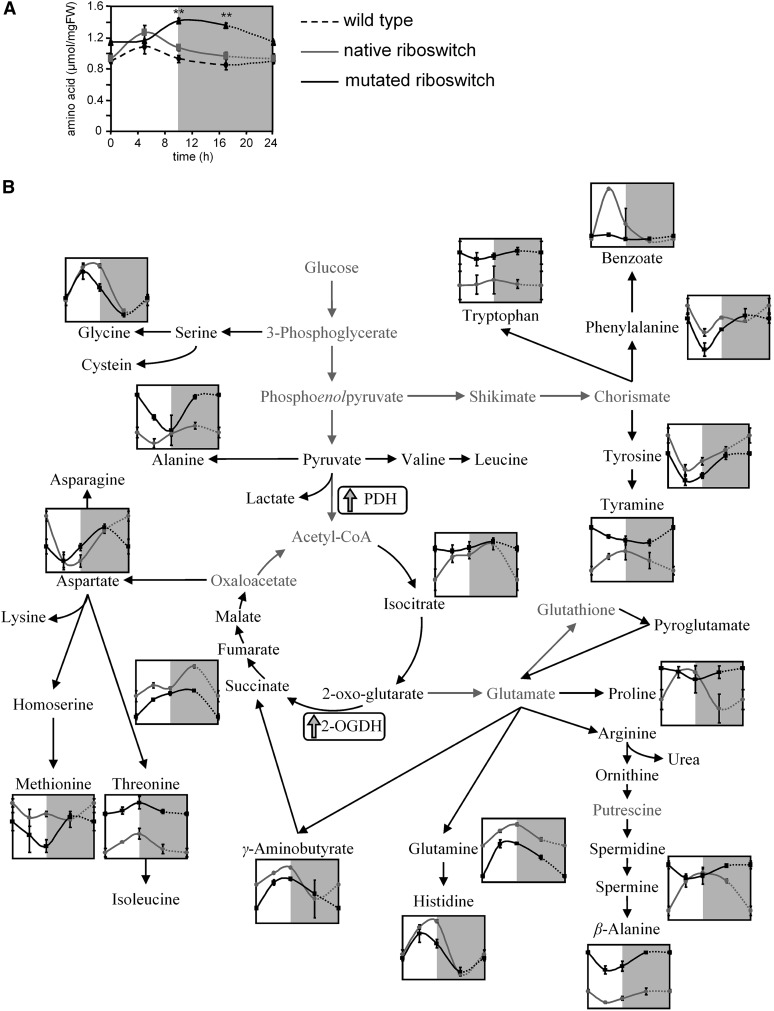
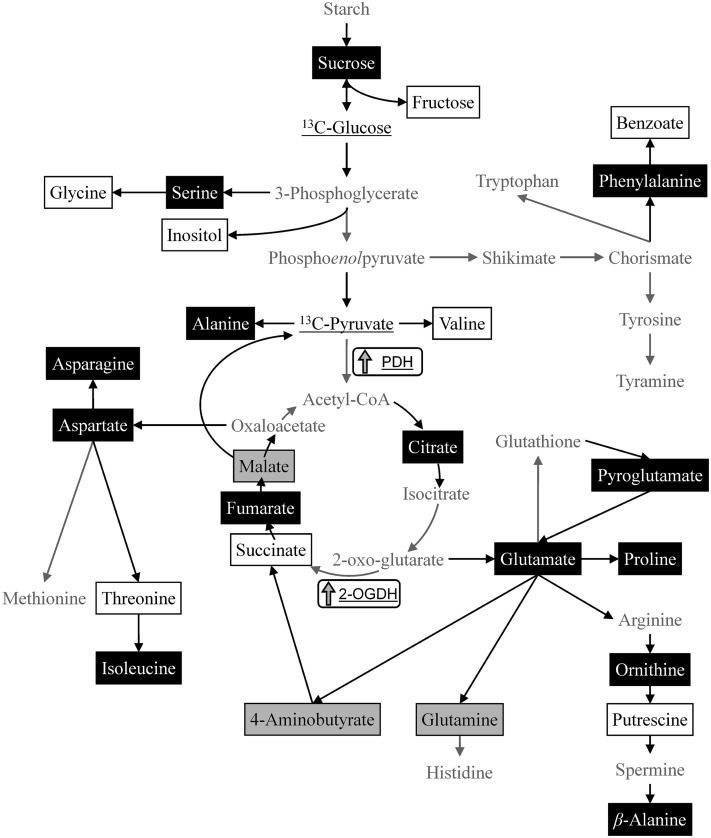
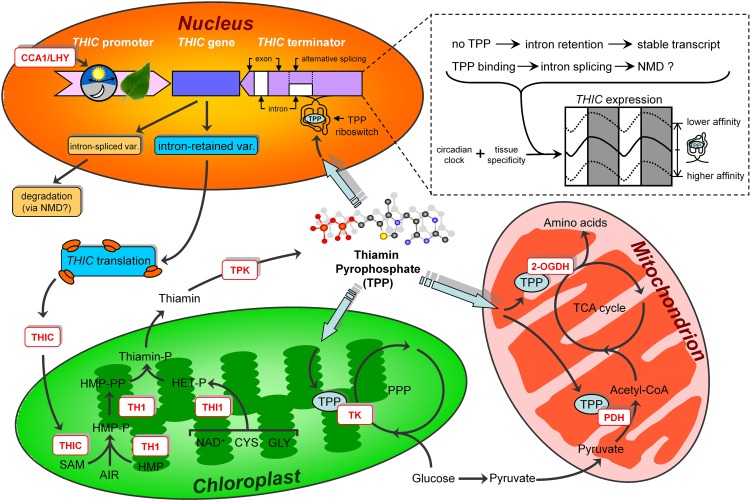
Riboswitches are natural RNA elements that posttranscriptionally regulate gene expression by binding small molecules and thereby autonomously control intracellular levels of these metabolites. Although riboswitch-based mechanisms have been examined extensively, the integration of their activity with global physiology and metabolism has been largely overlooked. Here, we explored the regulation of thiamin biosynthesis and the consequences of thiamin pyrophosphate riboswitch deficiency on metabolism in Arabidopsis thaliana. Our results show that thiamin biosynthesis is largely regulated by the circadian clock via the activity of the THIAMIN C SYNTHASE (THIC) promoter, while the riboswitch located at the 3' untranslated region of this gene controls overall thiamin biosynthesis. Surprisingly, the results also indicate that the rate of thiamin biosynthesis directs the activity of thiamin-requiring enzymes and consecutively determines the rate of carbohydrate oxidation via the tricarboxylic acid cycle and pentose-phosphate pathway. Our model suggests that in Arabidopsis, the THIC promoter and the thiamin-pyrophosphate riboswitch act simultaneously to tightly regulate thiamin biosynthesis in a circadian manner and consequently sense and control vital points of core cellular metabolism.
Figures
References
-
- Ajjawi I., Rodriguez Milla M.A., Cushman J., Shintani D.K. (2007b). Thiamin pyrophosphokinase is required for thiamin cofactor activation in Arabidopsis. Plant Mol. Biol. 65: 151–162 - PubMed
-
- Ajjawi I., Tsegaye Y., Shintani D. (2007a). Determination of the genetic, molecular, and biochemical basis of the Arabidopsis thaliana thiamin auxotroph th1. Arch. Biochem. Biophys. 459: 107–114 - PubMed
-
- Alabadí D., Oyama T., Yanovsky M.J., Harmon F.G., Más P., Kay S.A. (2001). Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 293: 880–883 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
