

Novel sexual-cycle-specific gene silencing in Aspergillus nidulans
- PMID: 23341415
- PMCID: PMC3606093
- DOI: 10.1534/genetics.112.147546
Novel sexual-cycle-specific gene silencing in Aspergillus nidulans
Abstract
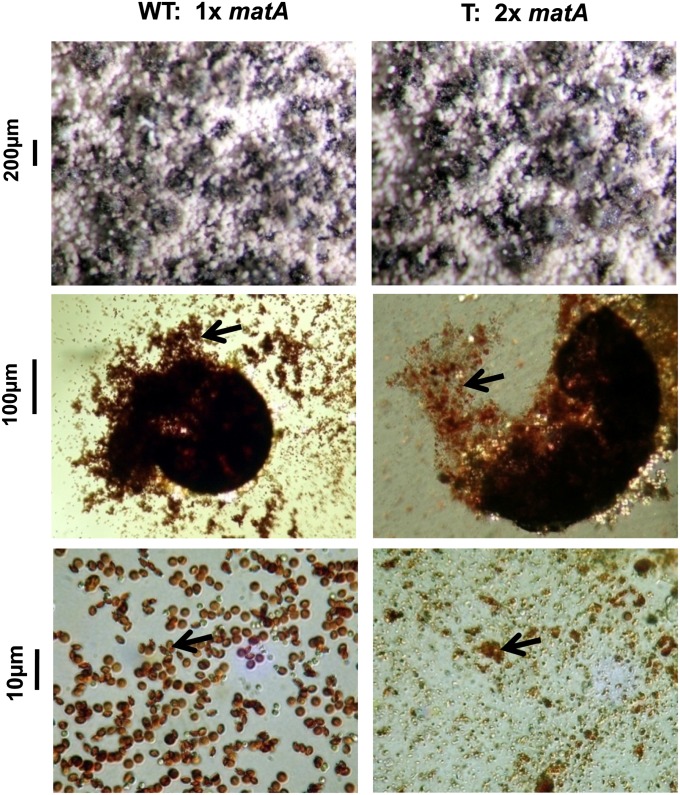
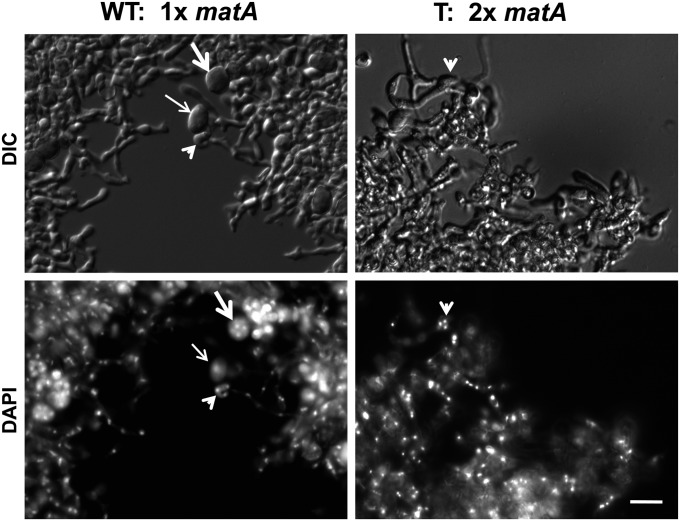
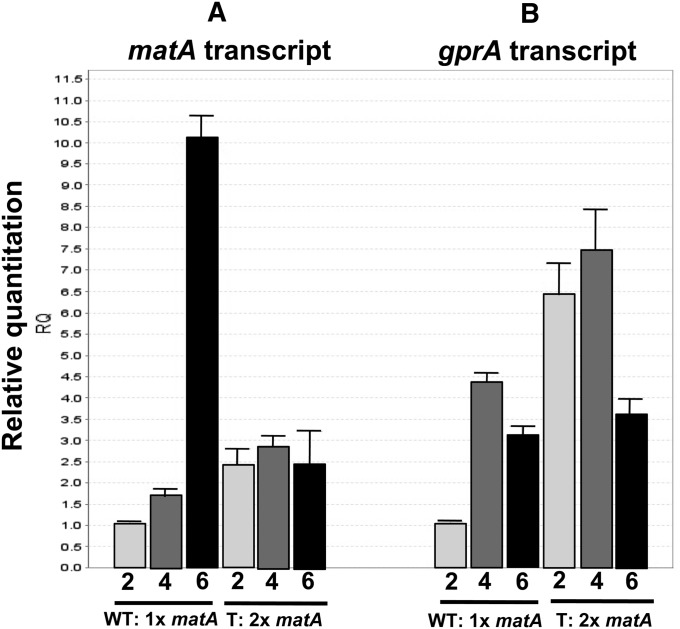
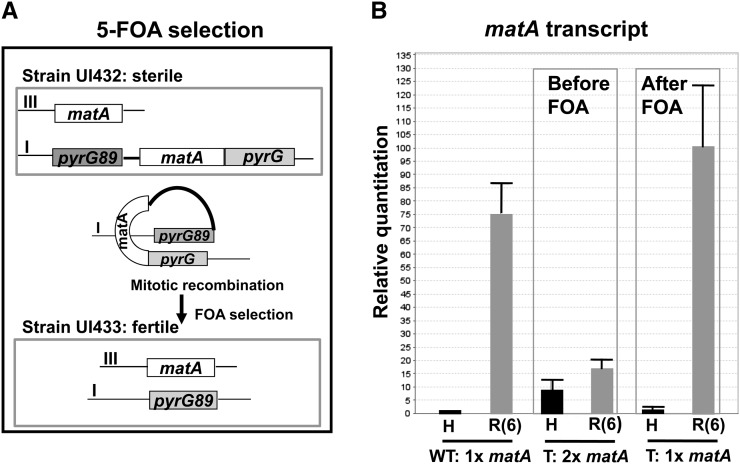
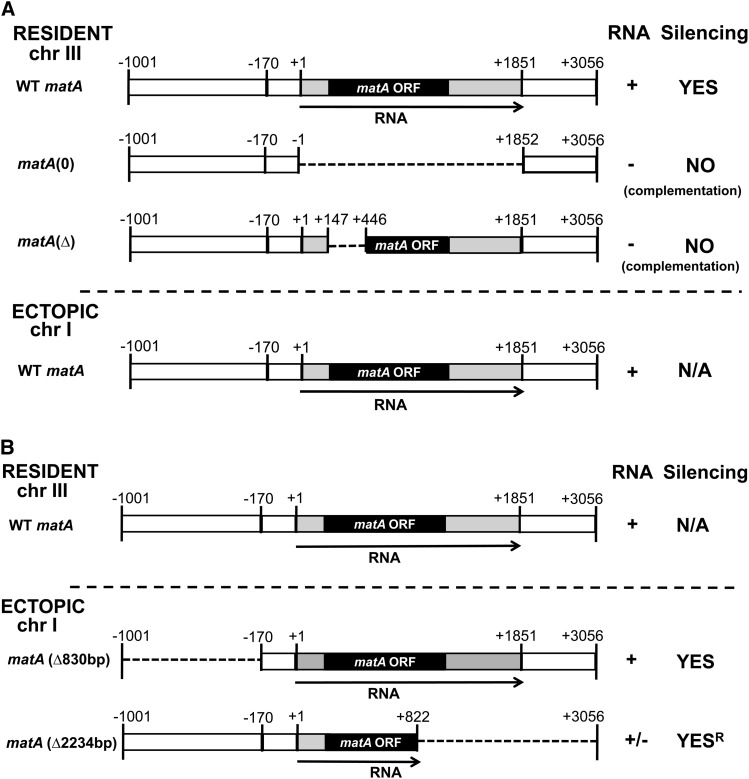
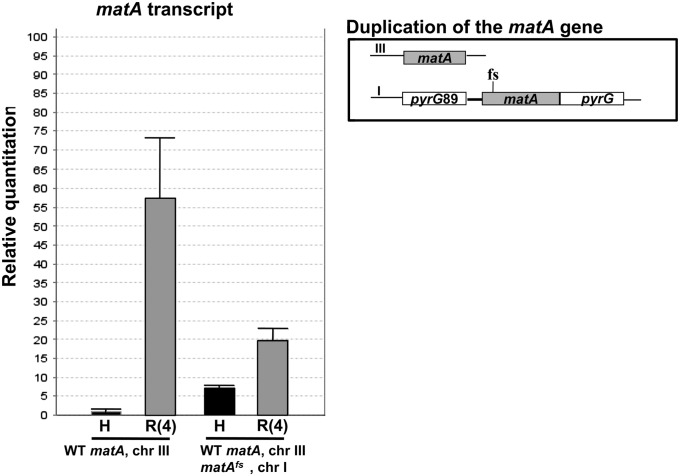
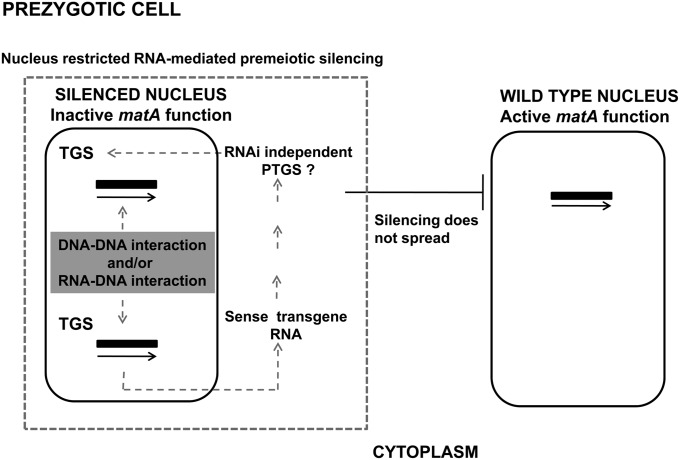
We report a novel sexual-cycle-specific gene-silencing system in the genetic model Aspergillus nidulans. Duplication of the mating type matA(HMG) gene in this haploid organism triggers Mat-induced silencing (MatIS) of both endogenous and transgenic matA genes, eliminates function of the encoded SRY structural ortholog, and results in formation of barren fruiting bodies. MatIS is spatiotemporally restricted to the prezygotic stage of the sexual cycle and does not interfere with vegetative growth, asexual reproduction, differentiation of early sexual tissues, or fruiting body development. MatIS is reversible upon deletion of the matA transgene. In contrast to other sex-specific silencing phenomena, MatIS silencing has nearly 100% efficiency and appears to be independent of homologous duplicated DNA segments. Remarkably, transgene-derived matA RNA might be sufficient to induce MatIS. A unique feature of MatIS is that RNA-mediated silencing is RNA interference/Argonaute-independent and is restricted to the nucleus having the duplicated gene. The silencing phenomenon is recessive and does not spread between nuclei within the common cytoplasm of a multinucleate heterokaryon. Gene silencing induced by matA gene duplication emerges as a specific feature associated with matA(HMG) regulation during sexual development.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
