

Forskolin-inducible cAMP pathway negatively regulates T-cell proliferation by uncoupling the interleukin-2 receptor complex
- PMID: 23341462
- PMCID: PMC3591623
- DOI: 10.1074/jbc.M112.408765
Forskolin-inducible cAMP pathway negatively regulates T-cell proliferation by uncoupling the interleukin-2 receptor complex
Abstract
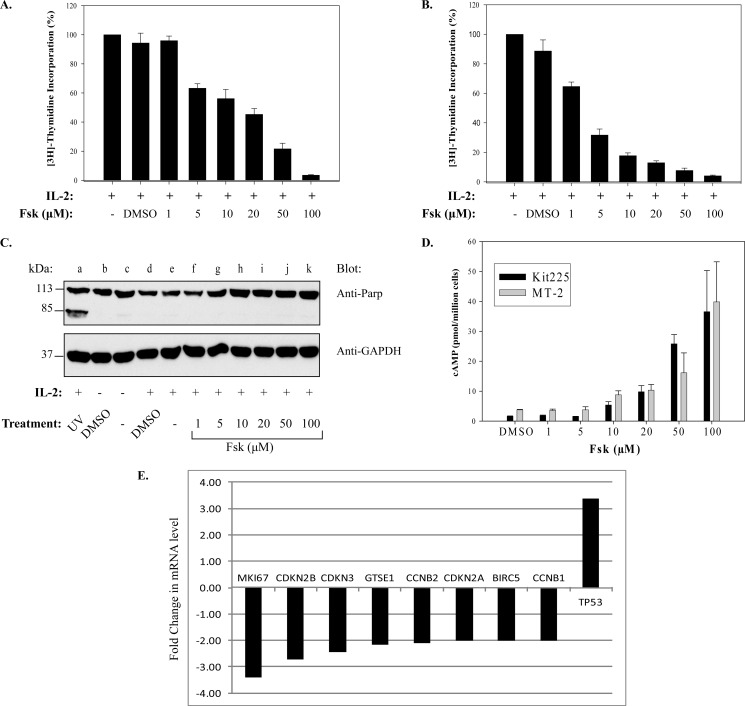
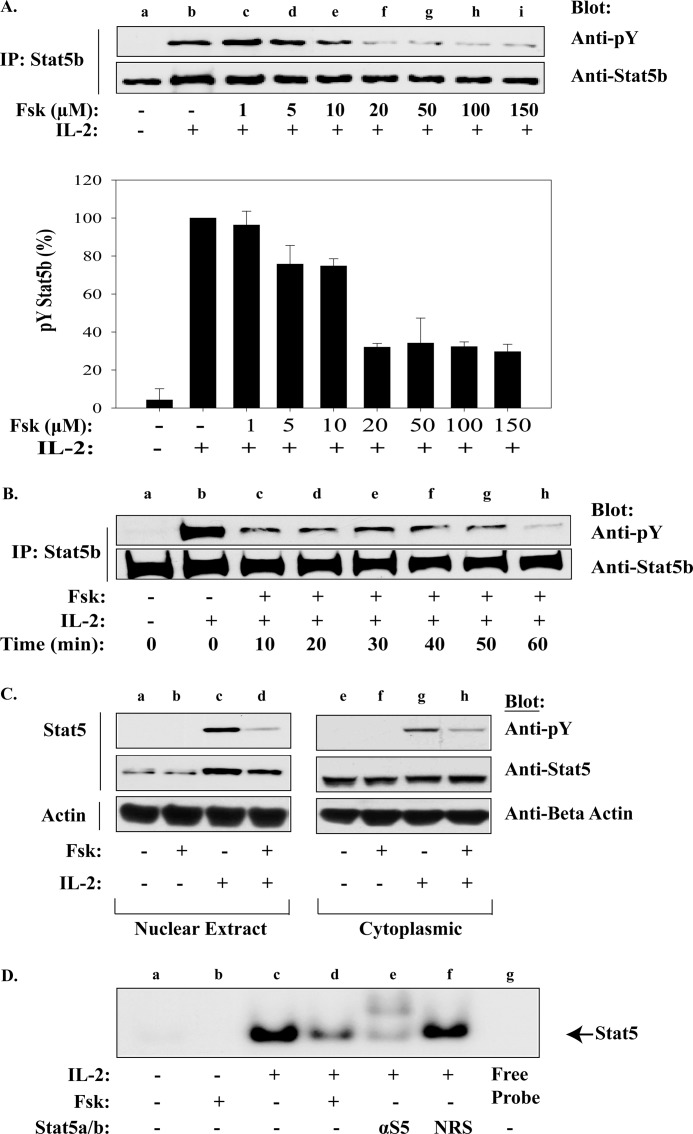
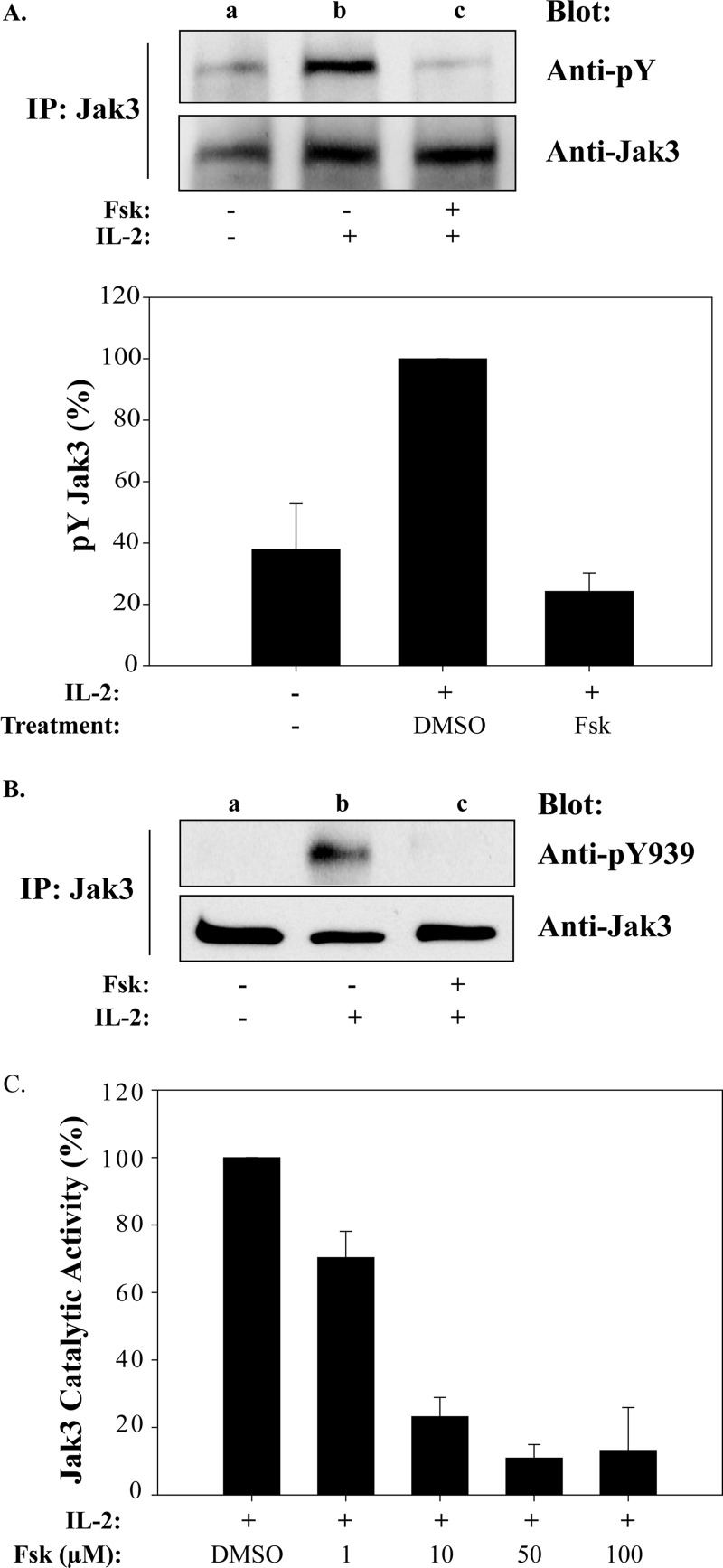
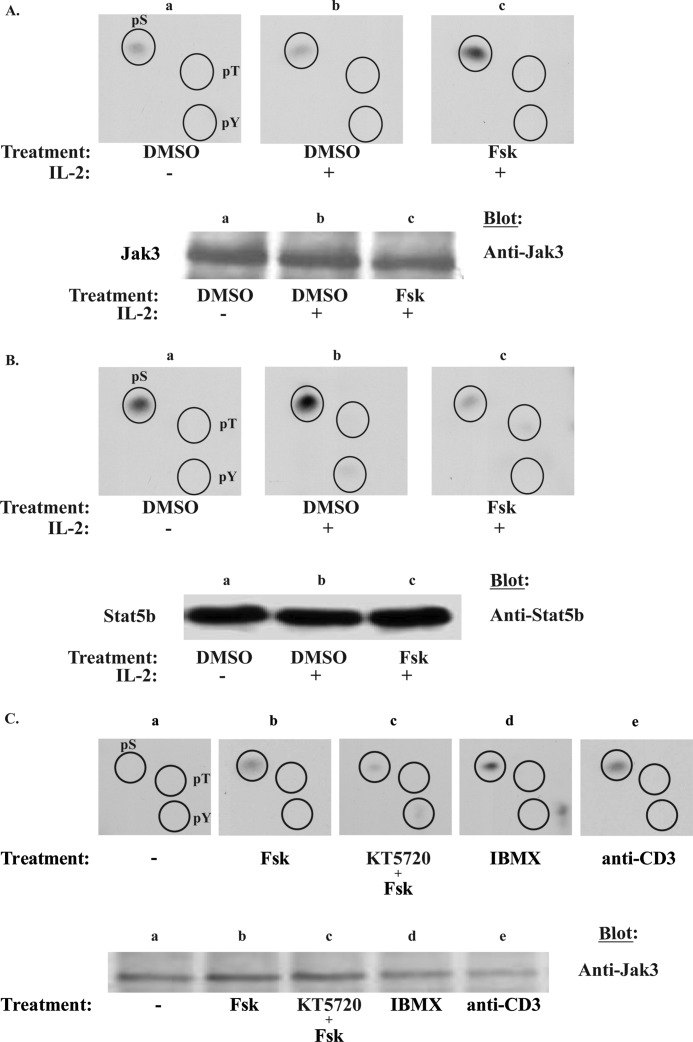
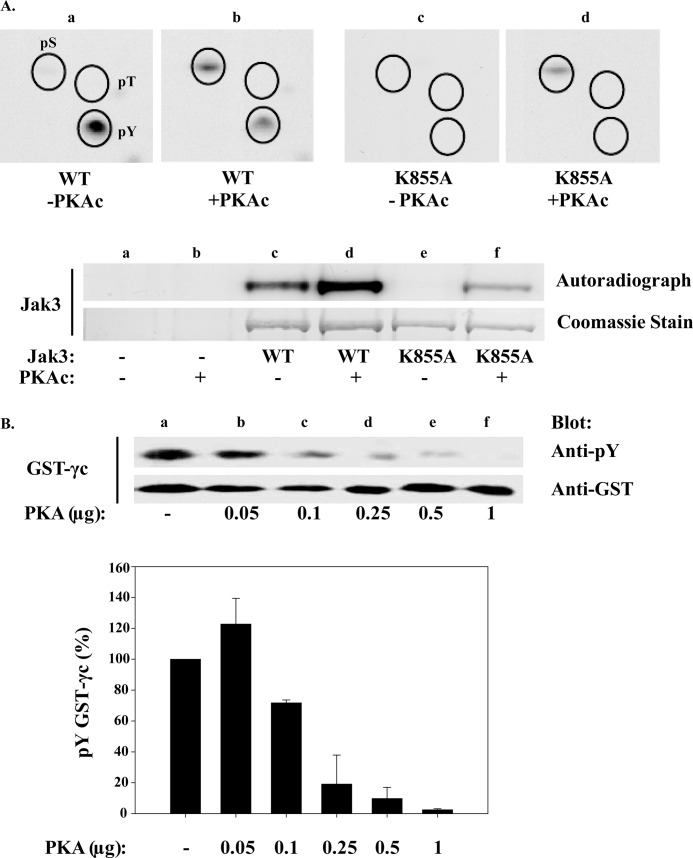
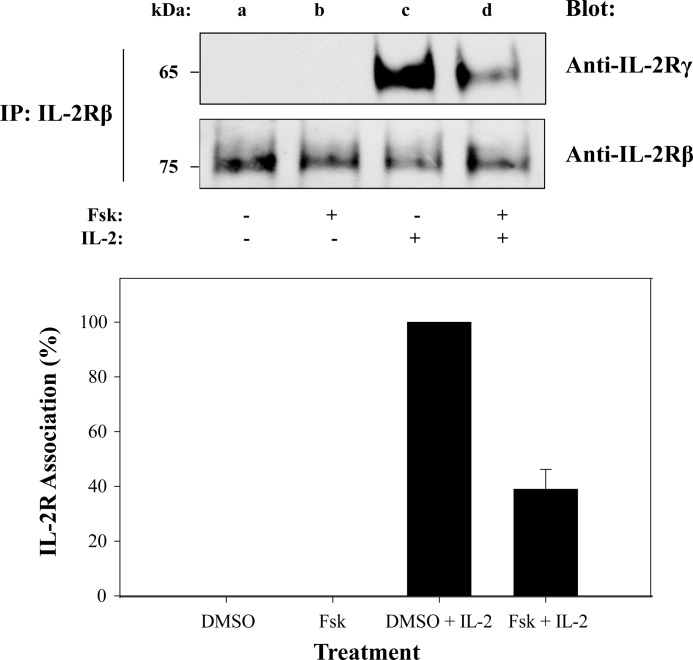
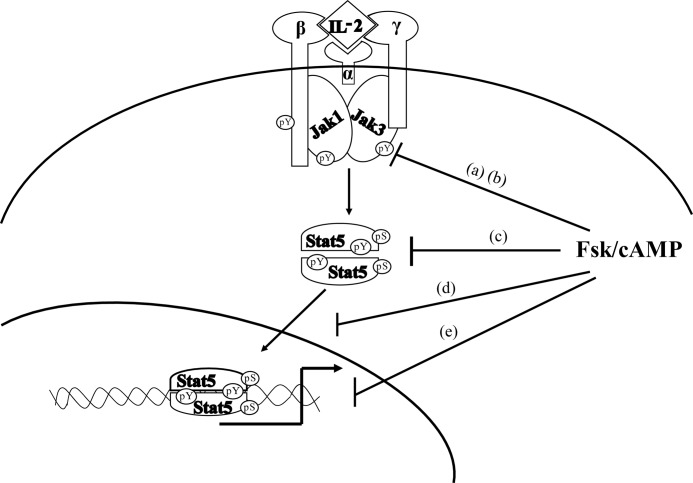
Cytokine-mediated regulation of T-cell activity involves a complex interplay between key signal transduction pathways. Determining how these signaling pathways cross-talk is essential to understanding T-cell function and dysfunction. In this work, we provide evidence that cross-talk exists between at least two signaling pathways: the Jak3/Stat5 and cAMP-mediated cascades. The adenylate cyclase activator forskolin (Fsk) significantly increased intracellular cAMP levels and reduced proliferation of the human T-cells via inhibition of cell cycle regulatory genes but did not induce apoptosis. To determine this inhibitory mechanism, effects of Fsk on IL-2 signaling was investigated. Fsk treatment of MT-2 and Kit 225 T-cells inhibited IL-2-induced Stat5a/b tyrosine and serine phosphorylation, nuclear translocation, and DNA binding activity. Fsk treatment also uncoupled IL-2 induced association of the IL-2Rβ and γc chain, consequently blocking Jak3 activation. Interestingly, phosphoamino acid analysis revealed that Fsk-treated cells resulted in elevated serine phosphorylation of Jak3 but not Stat5, suggesting that Fsk can negatively regulate Jak3 activity possibly mediated through PKA. Indeed, in vitro kinase assays and small molecule inhibition studies indicated that PKA can directly serine phosphorylate and functionally inactivate Jak3. Taken together, these findings suggest that Fsk activation of adenylate cyclase and PKA can negatively regulate IL-2 signaling at multiple levels that include IL-2R complex formation and Jak3/Stat5 activation.
Figures
Similar articles
-
Downregulation of JAK3 protein levels in T lymphocytes by prostaglandin E2 and other cyclic adenosine monophosphate-elevating agents: impact on interleukin-2 receptor signaling pathway.Blood. 1999 Apr 1;93(7):2308-18. Blood. 1999. PMID: 10090941
-
Protein phosphatase 2A regulates interleukin-2 receptor complex formation and JAK3/STAT5 activation.J Biol Chem. 2010 Feb 5;285(6):3582-3591. doi: 10.1074/jbc.M109.053843. Epub 2009 Nov 18. J Biol Chem. 2010. PMID: 19923221 Free PMC article.
-
Functional uncoupling of the Janus kinase 3-Stat5 pathway in malignant growth of human T cell leukemia virus type 1-transformed human T cells.J Immunol. 2000 Nov 1;165(9):5097-104. doi: 10.4049/jimmunol.165.9.5097. J Immunol. 2000. PMID: 11046040
-
The common γ-chain cytokine receptor: tricks-and-treats for T cells.Cell Mol Life Sci. 2016 Jan;73(2):253-69. doi: 10.1007/s00018-015-2062-4. Epub 2015 Oct 14. Cell Mol Life Sci. 2016. PMID: 26468051 Free PMC article. Review.
-
Signaling and Function of Interleukin-2 in T Lymphocytes.Annu Rev Immunol. 2018 Apr 26;36:411-433. doi: 10.1146/annurev-immunol-042617-053352. Annu Rev Immunol. 2018. PMID: 29677473 Free PMC article. Review.
Cited by
-
Pharmacological inhibition of hematopoietic progenitor kinase 1 positively regulates T-cell function.PLoS One. 2020 Dec 3;15(12):e0243145. doi: 10.1371/journal.pone.0243145. eCollection 2020. PLoS One. 2020. PMID: 33270695 Free PMC article.
-
Chemical reprogramming ameliorates cellular hallmarks of aging and extends lifespan.EMBO Mol Med. 2025 Aug;17(8):2071-2094. doi: 10.1038/s44321-025-00265-9. Epub 2025 Jun 30. EMBO Mol Med. 2025. PMID: 40588563 Free PMC article.
-
Elevation of cAMP Levels Inhibits Doxorubicin-Induced Apoptosis in Pre- B ALL NALM- 6 Cells Through Induction of BAD Phosphorylation and Inhibition of P53 Accumulation.Int J Mol Cell Med. 2015 Spring;4(2):94-102. Int J Mol Cell Med. 2015. PMID: 26261798 Free PMC article.
-
Molecular Mechanisms for cAMP-Mediated Immunoregulation in T cells - Role of Anchored Protein Kinase A Signaling Units.Front Immunol. 2016 Jun 8;7:222. doi: 10.3389/fimmu.2016.00222. eCollection 2016. Front Immunol. 2016. PMID: 27375620 Free PMC article. Review.
-
Extracellular metallothionein: An alarmin regulating lymphocyte gene expression, cell signaling, and immune function.Cell Stress Chaperones. 2025 Jul 31;30(5):100106. doi: 10.1016/j.cstres.2025.100106. Online ahead of print. Cell Stress Chaperones. 2025. PMID: 40752583 Free PMC article.
References
-
- Leonard W. J. (1996) STATs and cytokine specificity. Nat. Med. 2, 968–969 - PubMed
-
- Weaver A. M., Silva C. M. (2006) Modulation of signal transducer and activator of transcription 5b activity in breast cancer cells by mutation of tyrosines within the transactivation domain. Mol. Endocrinol. 20, 2392–2405 - PubMed
-
- Lin J. X., Mietz J., Modi W. S., John S., Leonard W. J. (1996) Cloning of human Stat5B. Reconstitution of interleukin-2-induced Stat5A and Stat5B DNA binding activity in COS-7 cells. J. Biol. Chem. 271, 10738–10744 - PubMed
-
- Lin J. X., Leonard W. J. (2000) The role of Stat5a and Stat5b in signaling by IL-2 family cytokines. Oncogene 19, 2566–2576 - PubMed
-
- Park S. H., Yamashita H., Rui H., Waxman D. J. (2001) Serine phosphorylation of GH-activated signal transducer and activator of transcription 5a (STAT5a) and STAT5b: impact on STAT5 transcriptional activity. Mol. Endocrinol. 15, 2157–2171 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
