

Activated invariant NKT cells control central nervous system autoimmunity in a mechanism that involves myeloid-derived suppressor cells
- PMID: 23345328
- PMCID: PMC3577977
- DOI: 10.4049/jimmunol.1201718
Activated invariant NKT cells control central nervous system autoimmunity in a mechanism that involves myeloid-derived suppressor cells
Abstract
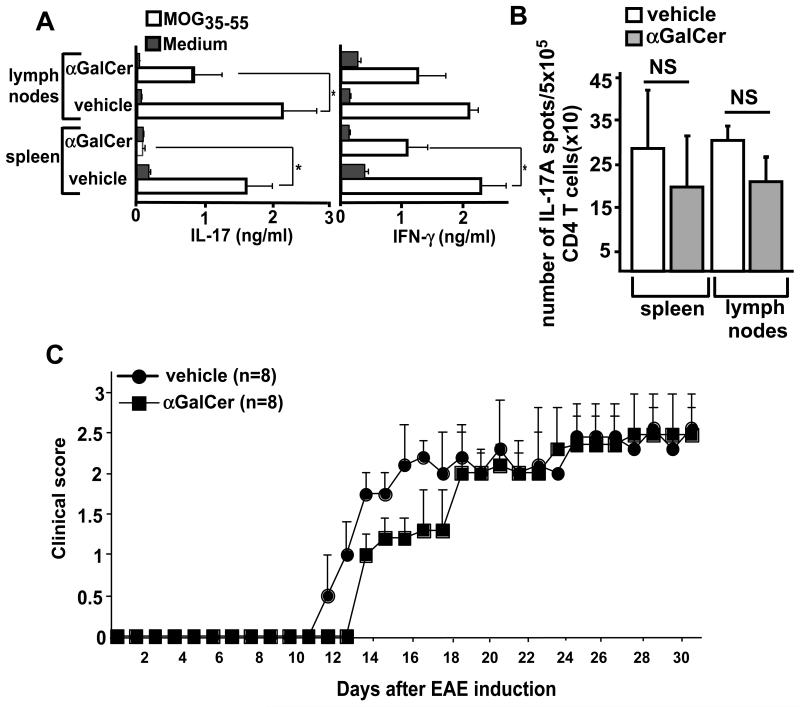
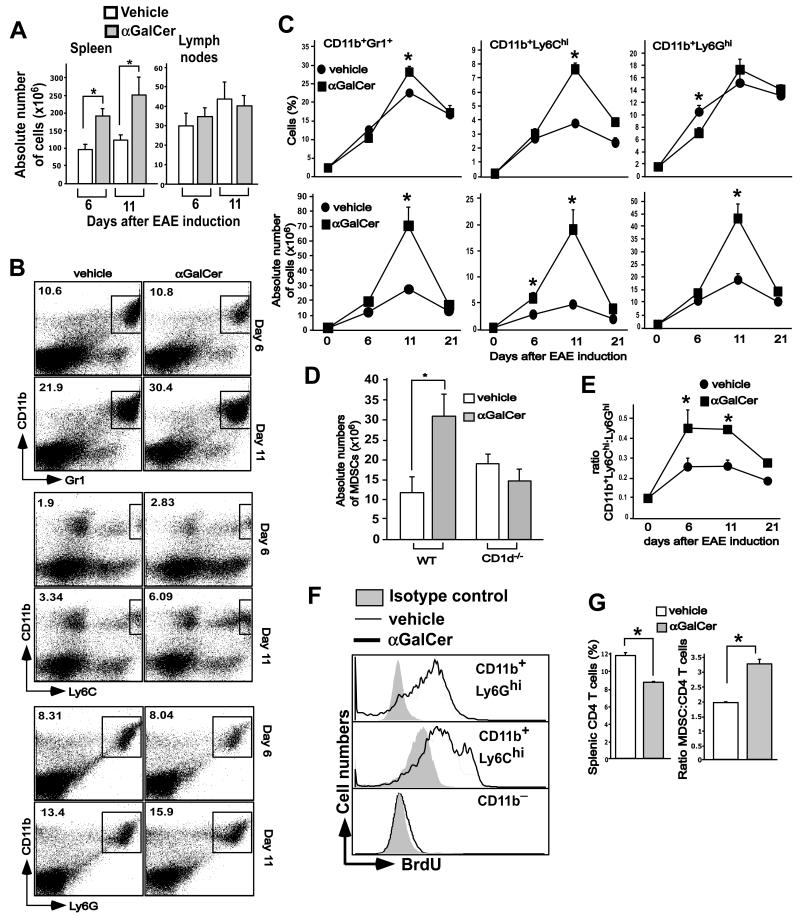
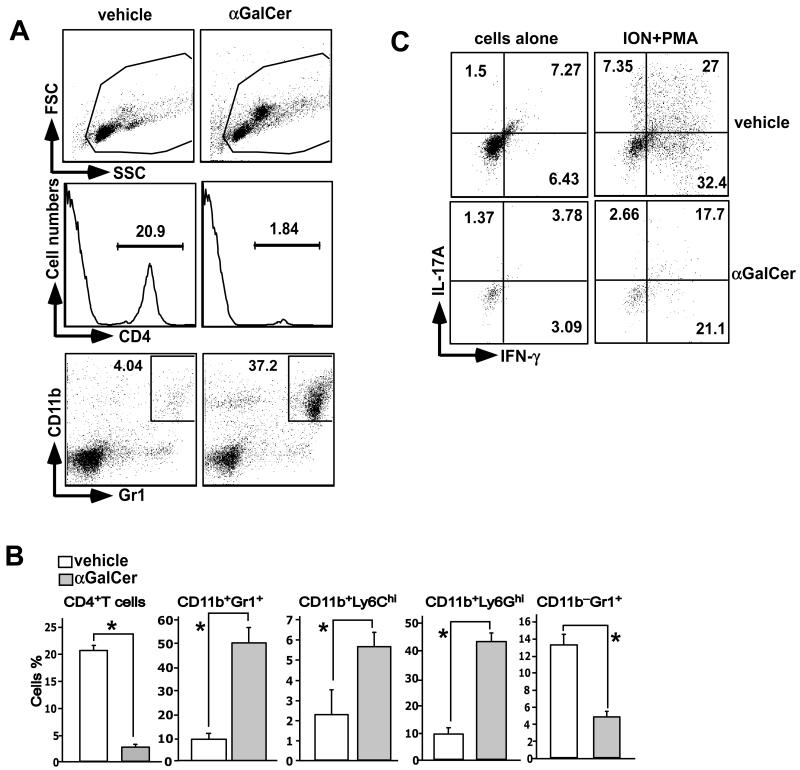
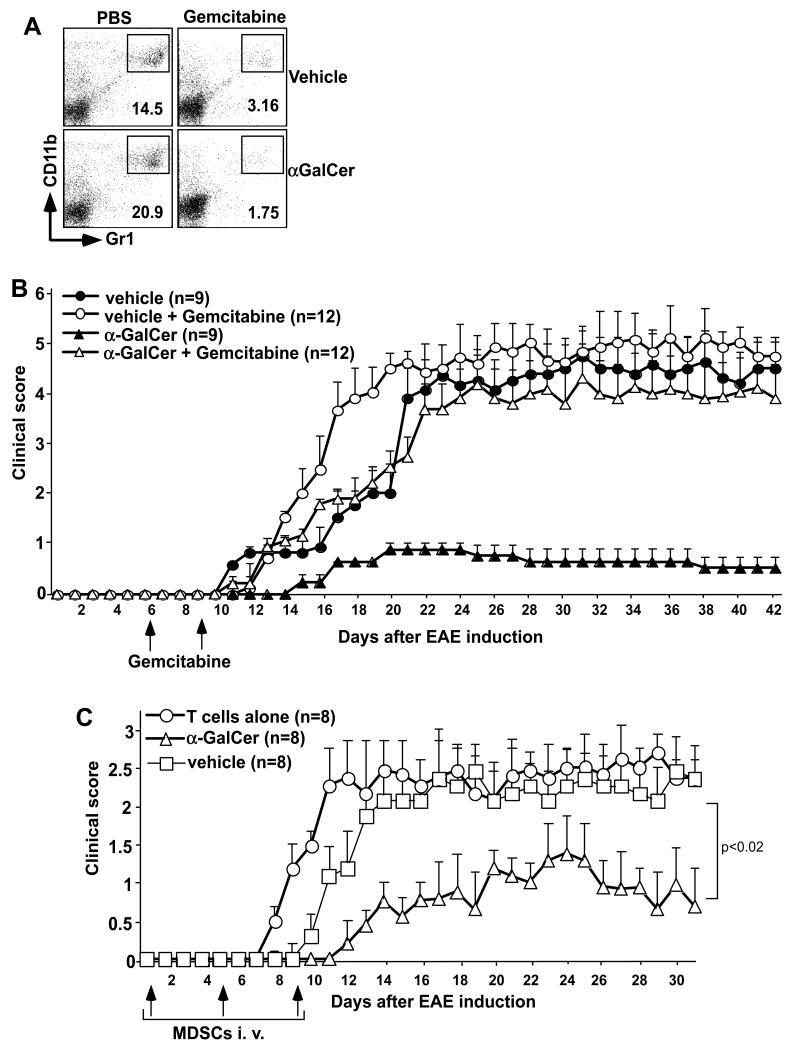
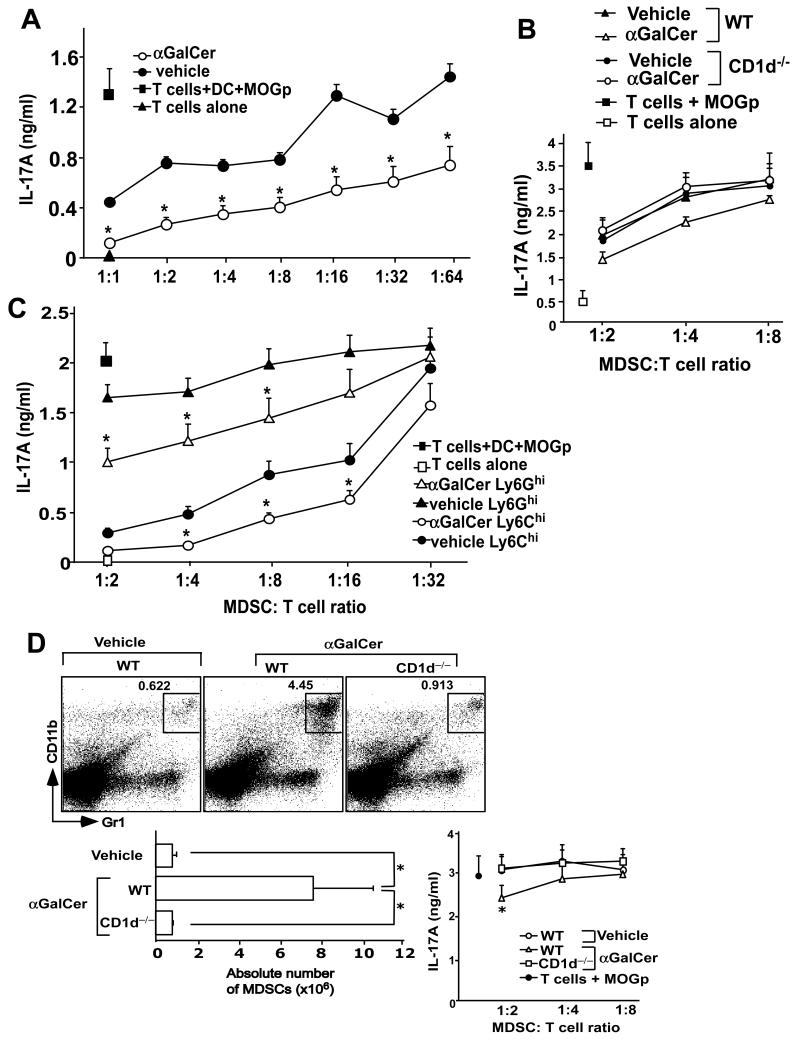
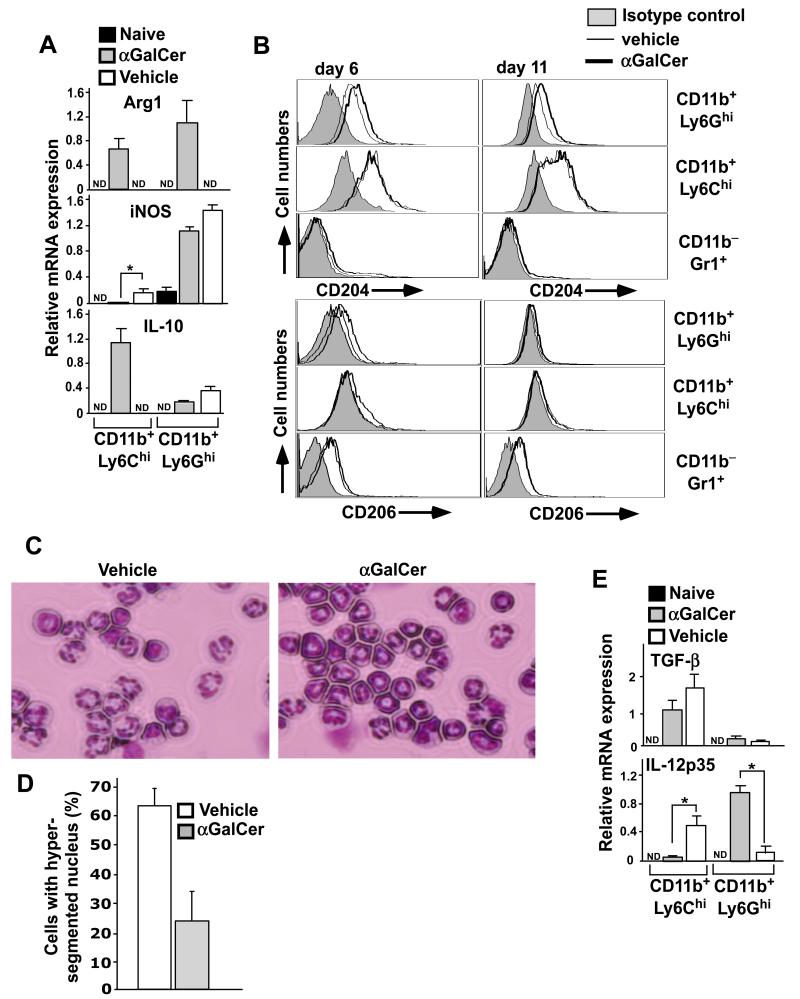
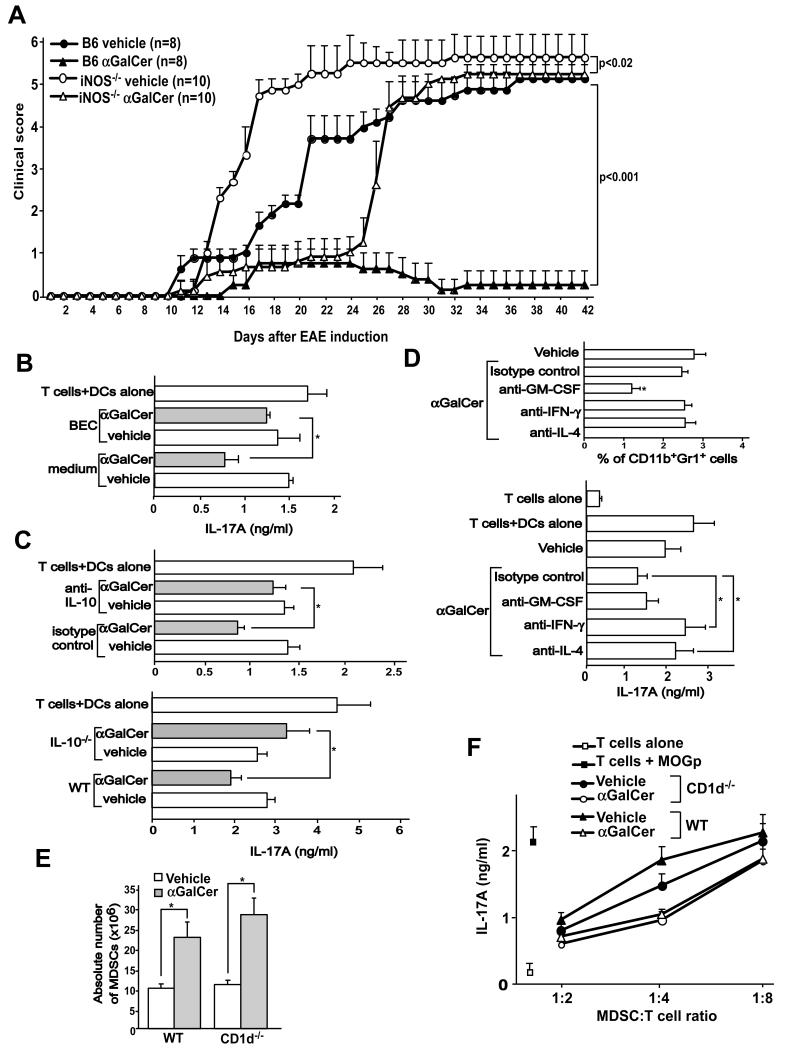
Invariant NKT (iNKT) cells are a subset of T lymphocytes that recognize glycolipid Ags presented by the MHC class I-related protein CD1d. Activation of iNKT cells with glycolipid Ags, such as the marine sponge-derived reagent α-galactosylceramide (α-GalCer), results in the rapid production of a variety of cytokines and activation of many other immune cell types. These immunomodulatory properties of iNKT cells have been exploited for the development of immunotherapies against a variety of autoimmune and inflammatory diseases, but mechanisms by which activated iNKT cells confer disease protection have remained incompletely understood. In this study, we demonstrate that glycolipid-activated iNKT cells cooperate with myeloid-derived suppressor cells (MDSCs) in protecting mice against the development of experimental autoimmune encephalomyelitis (EAE) in mice, an animal model for multiple sclerosis. We show that α-GalCer induced the expansion and immunosuppressive activities of MDSCs in the spleen of mice induced for development of EAE. Disease protection in these animals also correlated with recruitment of MDSCs to the CNS. Depletion of MDSCs abrogated the protective effects of α-GalCer against EAE and, conversely, adoptive transfer of MDSCs from α-GalCer-treated mice ameliorated passive EAE induced in recipient animals. The cytokines GM-CSF, IL-4, and IFN-γ, produced by activated iNKT cells, and inducible NO synthase, arginase-1, and IL-10 produced by MDSCs, contributed to these effects. Our findings have revealed cooperative immunosuppressive interactions between iNKT cells and MDSCs that might be exploited for the development of improved immunotherapies for multiple sclerosis and other autoimmune and inflammatory diseases.
Figures
References
-
- Bendelac A, Savage PB, Teyton L. The biology of NKT cells. Annu Rev Immunol. 2007;25:297–336. - PubMed
-
- Taniguchi M, Harada M, Kojo S, Nakayama T, Wakao H. The regulatory role of Valpha14 NKT cells in innate and acquired immune response. Annu Rev Immunol. 2003;21:483–513. - PubMed
-
- Kronenberg M. Toward an understanding of NKT cell biology: progress and paradoxes. Annu Rev Immunol. 2005;23:877–900. - PubMed
-
- Brigl M, Brenner MB. CD1: antigen presentation and T cell function. Annu Rev Immunol. 2004;22:817–890. - PubMed
-
- Parekh VV, Lalani S, Van Kaer L. The in vivo response of invariant natural killer T cells to glycolipid antigens. Int Rev Immunol. 2007;26:31–48. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
