

Rearrangements within human spliceosomes captured after exon ligation
- PMID: 23345524
- PMCID: PMC3677250
- DOI: 10.1261/rna.034223.112
Rearrangements within human spliceosomes captured after exon ligation
Abstract
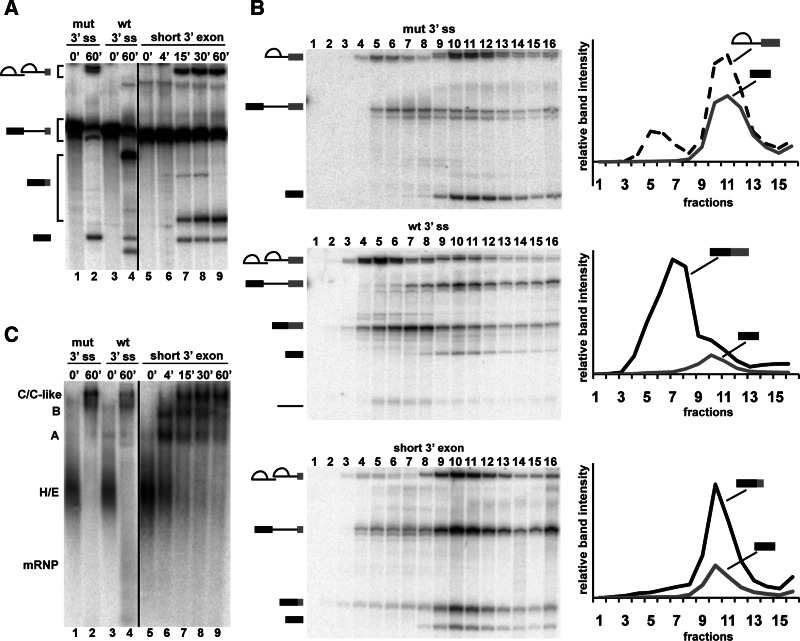
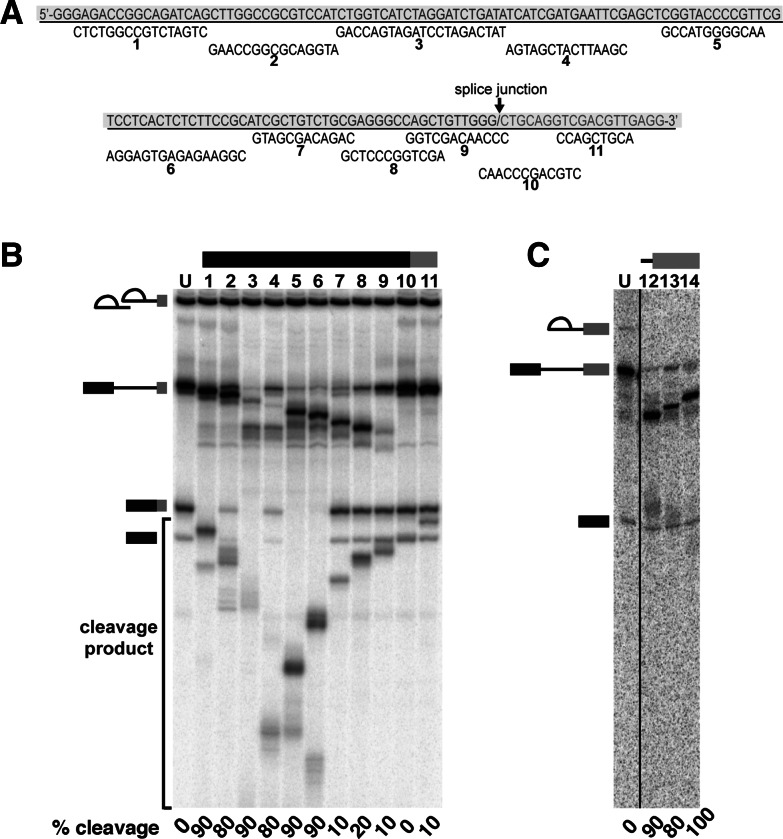
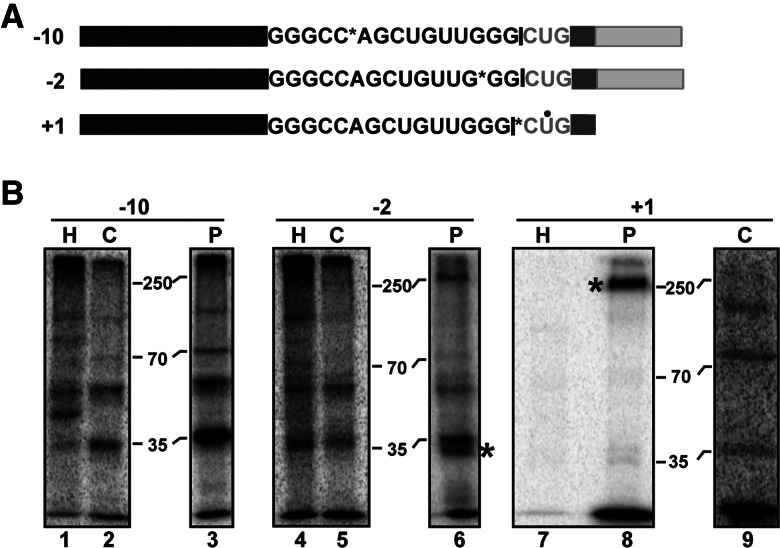
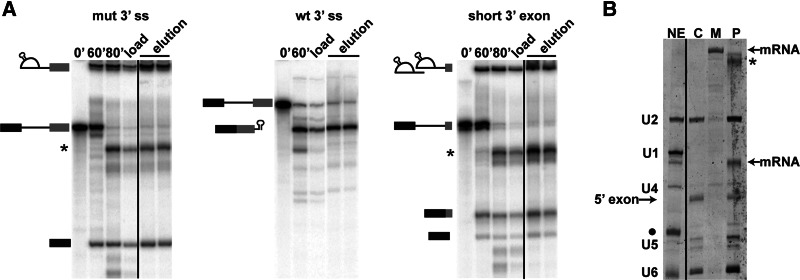
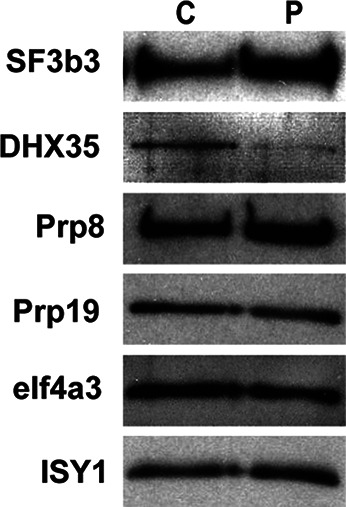
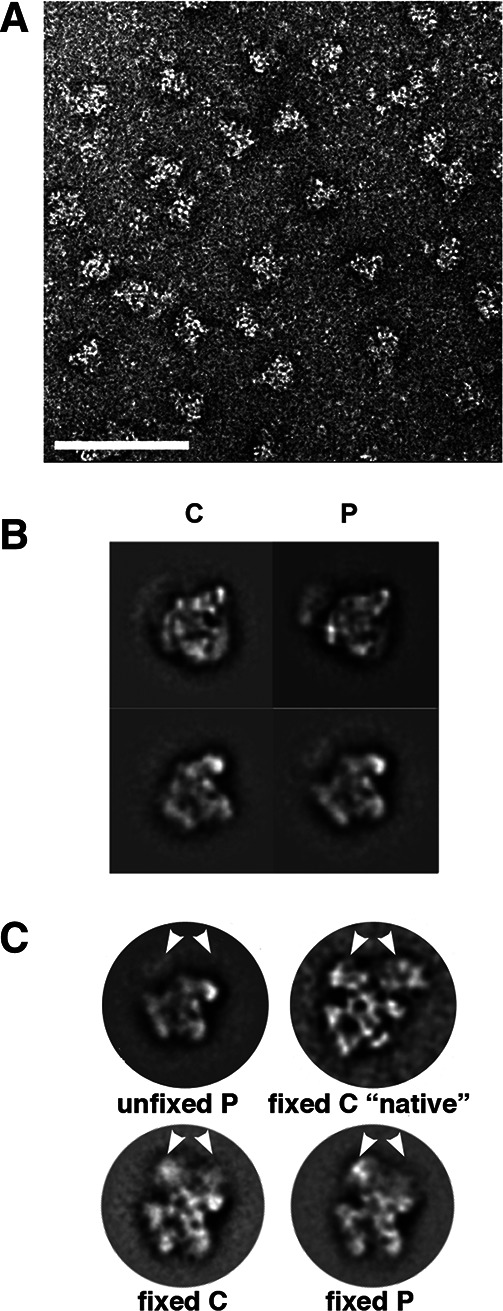
In spliceosomes, dynamic RNA/RNA and RNA/protein interactions position the pre-mRNA substrate for the two chemical steps of splicing. Not all of these interactions have been characterized, in part because it has not been possible to arrest the complex at clearly defined states relative to chemistry. Previously, it was shown in yeast that the DEAD/H-box protein Prp22 requires an extended 3' exon to promote mRNA release from the spliceosome following second-step chemistry. In line with that observation, we find that shortening the 3' exon blocks cleaved lariat intron and mRNA release in human splicing extracts, which allowed us to stall human spliceosomes in a new post-catalytic complex (P complex). In comparison to C complex, which is blocked at a point following first-step chemistry, we detect specific differences in RNA substrate interactions near the splice sites. These differences include extended protection across the exon junction and changes in protein crosslinks to specific sites in the 5' and 3' exons. Using selective reaction monitoring (SRM) mass spectrometry, we quantitatively compared P and C complex proteins and observed enrichment of SF3b components and loss of the putative RNA-dependent ATPase DHX35. Electron microscopy revealed similar structural features for both complexes. Notably, additional density is present when complexes are chemically fixed, which reconciles our results with previously reported C complex structures. Our ability to compare human spliceosomes before and after second-step chemistry has opened a new window to rearrangements near the active site of spliceosomes, which may play roles in exon ligation and mRNA release.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- RR015804/RR/NCRR NIH HHS/United States
- T32GM08646/GM/NIGMS NIH HHS/United States
- RR001614/RR/NCRR NIH HHS/United States
- RR019934/RR/NCRR NIH HHS/United States
- T32 GM008646/GM/NIGMS NIH HHS/United States
- R01 GM072649/GM/NIGMS NIH HHS/United States
- S10 RR019934/RR/NCRR NIH HHS/United States
- R25 GM058903/GM/NIGMS NIH HHS/United States
- P41 RR001614/RR/NCRR NIH HHS/United States
- R01GM72649/GM/NIGMS NIH HHS/United States
- S10 RR015804/RR/NCRR NIH HHS/United States
- R25GM058903/GM/NIGMS NIH HHS/United States
- P41 GM103481/GM/NIGMS NIH HHS/United States
- P41GM103481/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases