

Histone H4 lysine 20 methylation: key player in epigenetic regulation of genomic integrity
- PMID: 23345616
- PMCID: PMC3597678
- DOI: 10.1093/nar/gkt012
Histone H4 lysine 20 methylation: key player in epigenetic regulation of genomic integrity
Abstract
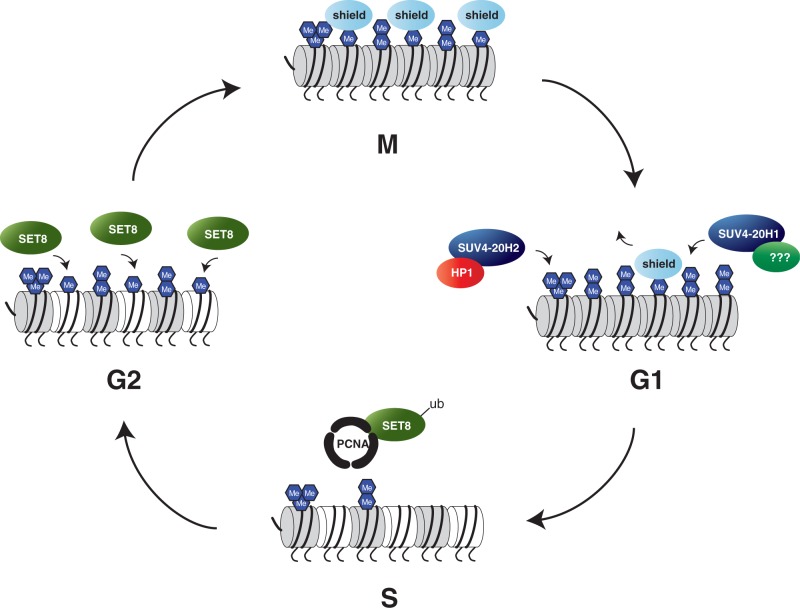
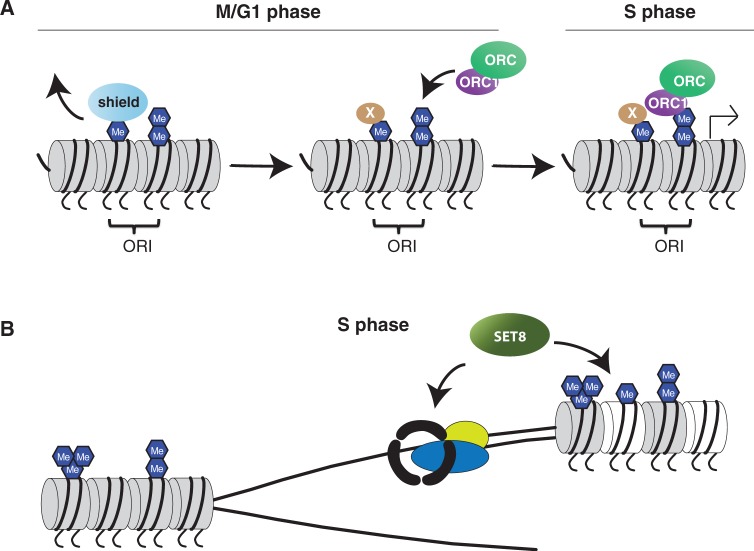
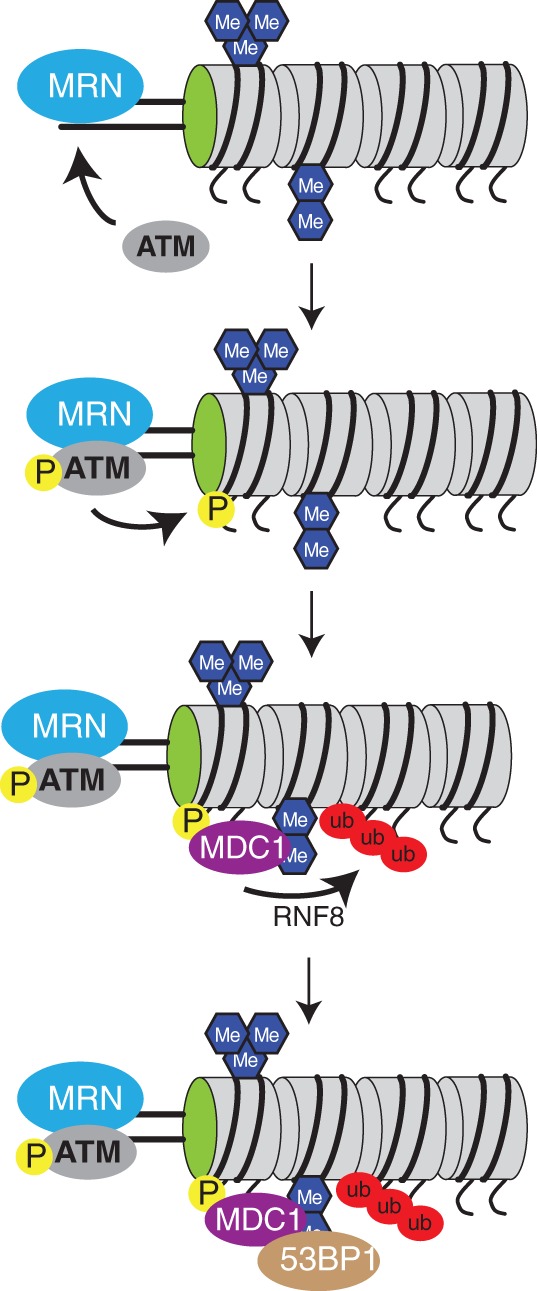
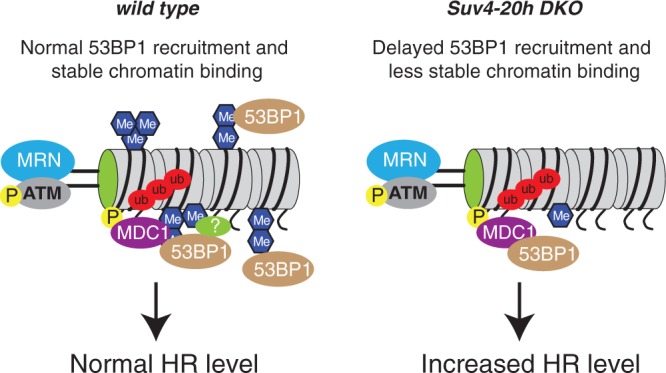
Maintenance of genomic integrity is essential to ensure normal organismal development and to prevent diseases such as cancer. Nuclear DNA is packaged into chromatin, and thus genome maintenance can be influenced by distinct chromatin environments. In particular, post-translational modifications of histones have emerged as key regulators of genomic integrity. Intense research during the past few years has revealed histone H4 lysine 20 methylation (H4K20me) as critically important for the biological processes that ensure genome integrity, such as DNA damage repair, DNA replication and chromatin compaction. The distinct H4K20 methylation states are mediated by SET8/PR-Set7 that catalyses monomethylation of H4K20, whereas SUV4-20H1 and SUV4-20H2 enzymes mediate further H4K20 methylation to H4K20me2 and H4K20me3. Disruption of these H4K20-specific histone methyltransferases leads to genomic instability, demonstrating the important functions of H4K20 methylation in genome maintenance. In this review, we explain molecular mechanisms underlying these defects and discuss novel ideas for furthering our understanding of genome maintenance in higher eukaryotes.
Figures
References
-
- Kouzarides T. Chromatin modifications and their function. Cell. 2007;128:693–705. - PubMed
-
- Lukas J, Lukas C, Bartek J. More than just a focus: the chromatin response to DNA damage and its role in genome integrity maintenance. Nat. Cell Biol. 2011;13:1161–1169. - PubMed
-
- Fang J, Feng Q, Ketel CS, Wang H, Cao R, Xia L, Erdjument-Bromage H, Tempst P, Simon JA, Zhang Y. Purification and functional characterization of SET8, a nucleosomal histone H4-lysine 20-specific methyltransferase. Curr. Biol. 2002;12:1086–1099. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
