

Genesis and regulatory wiring of retroelement-derived domesticated genes: a phylogenomic perspective
- PMID: 23348003
- PMCID: PMC3670739
- DOI: 10.1093/molbev/mst014
Genesis and regulatory wiring of retroelement-derived domesticated genes: a phylogenomic perspective
Abstract
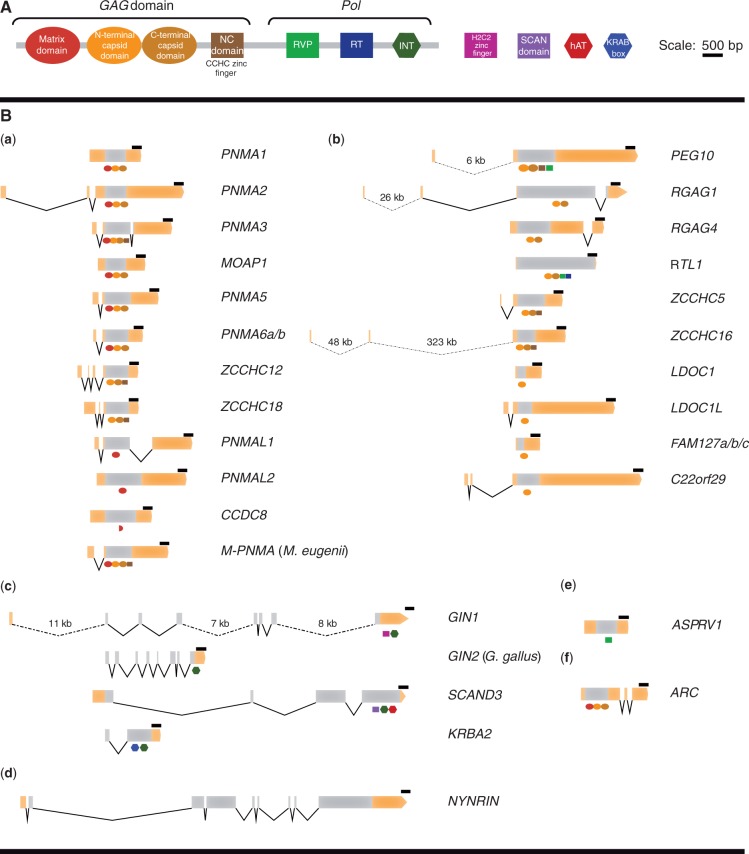
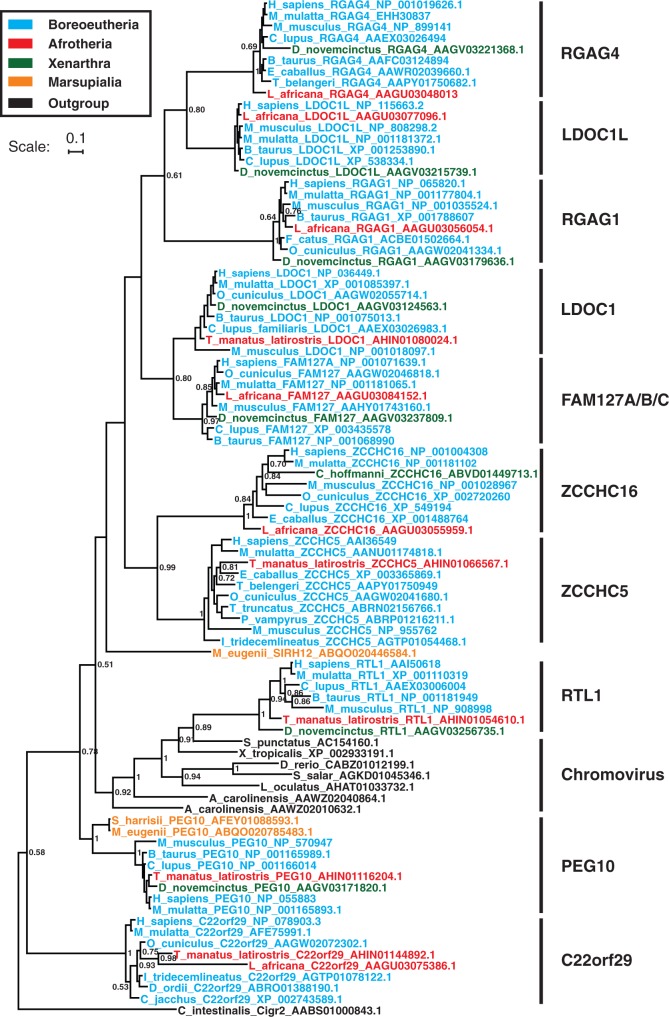
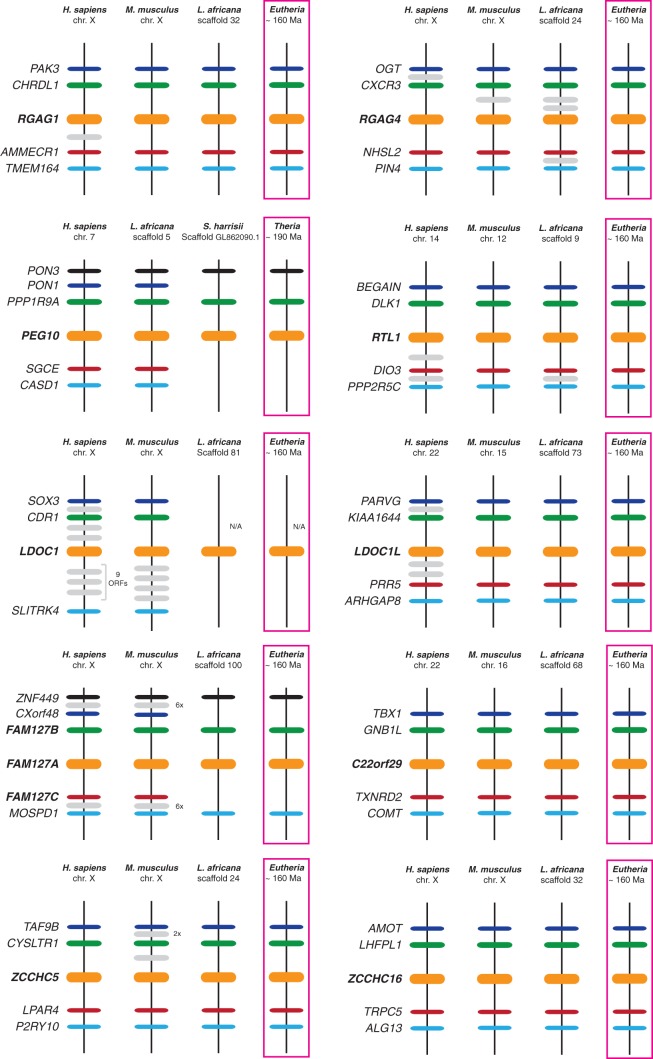
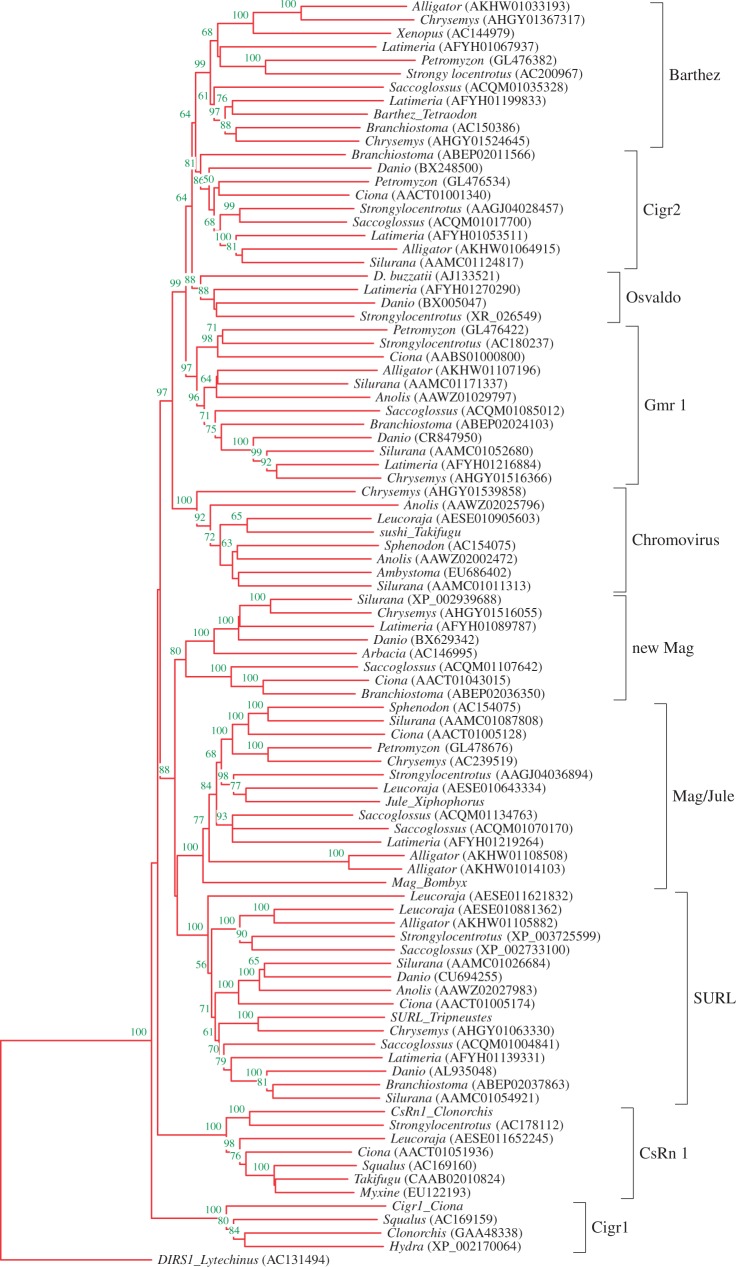
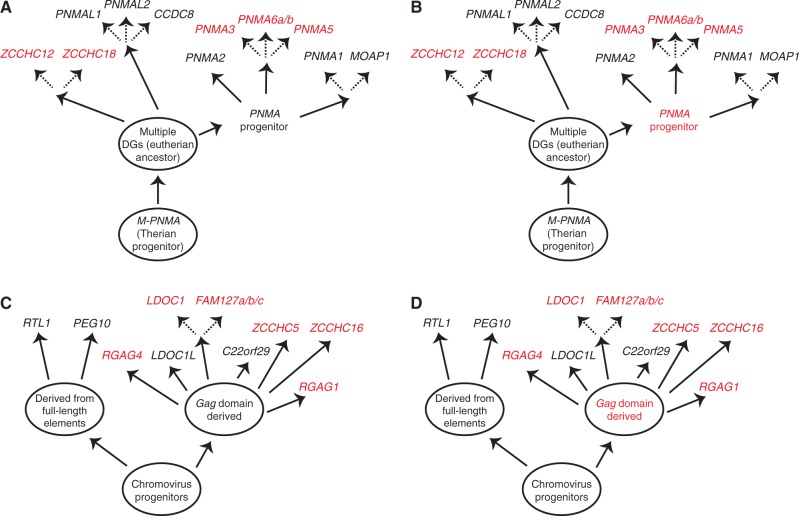
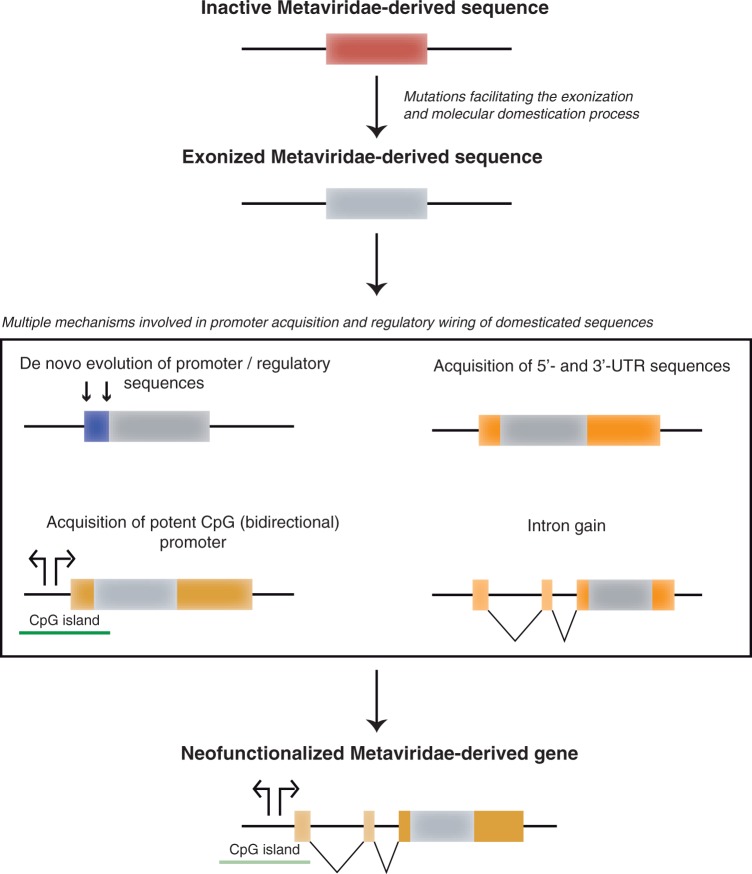
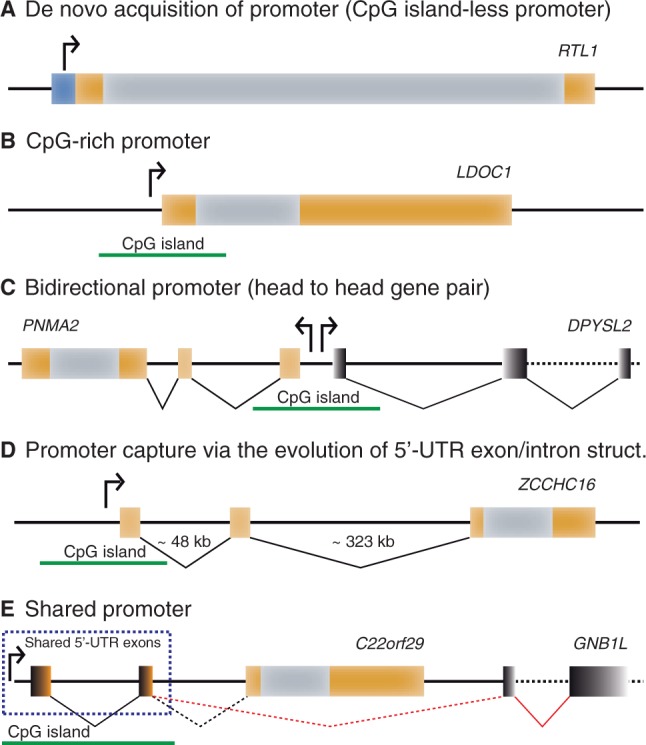
Molecular domestications of transposable elements have occurred repeatedly during the evolution of eukaryotes. Vertebrates, especially mammals, possess numerous single copy domesticated genes (DGs) that have originated from the intronless multicopy transposable elements. However, the origin and evolution of the retroelement-derived DGs (RDDGs) that originated from Metaviridae has been only partially elucidated, due to absence of genome data or to limited analysis of a single family of DGs. We traced the genesis and regulatory wiring of the Metaviridae-derived DGs through phylogenomic analysis, using whole-genome information from more than 90 chordate genomes. Phylogenomic analysis of these DGs in chordate genomes provided direct evidence that major diversification has occurred in the ancestor of placental mammals. Mammalian RDDGs have been shown to originate in several steps by independent domestication events and to diversify later by gene duplications. Analysis of syntenic loci has shown that diverse RDDGs and their chromosomal positions were fully established in the ancestor of placental mammals. By analysis of active Metaviridae lineages in amniotes, we have demonstrated that RDDGs originated from retroelement remains. The chromosomal gene movements of RDDGs were highly dynamic only in the ancestor of placental mammals. During the domestication process, de novo acquisition of regulatory regions is shown to be a prerequisite for the survival of the DGs. The origin and evolution of de novo acquired promoters and untranslated regions in diverse mammalian RDDGs have been explained by comparative analysis of orthologous gene loci. The origin of placental mammal-specific innovations and adaptations, such as placenta and newly evolved brain functions, was most probably connected to the regulatory wiring of DGs and their rapid fixation in the ancestor of placental mammals.
Figures
References
-
- Almeida LM, Silva IT, Silva WA, Jr, Castro JP, Riggs PK, Carareto CM, Amaral ME. The contribution of transposable elements to Bos taurus gene structure. Gene. 2007;390:180–189. - PubMed
-
- Andreassi C, Riccio A. To localize or not to localize: mRNA fate is in 3'UTR ends. Trends Cell Biol. 2009;19:465–474. - PubMed
-
- Benton MJ, Twitchett RJ. How to kill (almost) all life: the end-Permian extinction event. Trends Ecol Evol. 2003;18:358–365.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
