

Metabotropic glutamate receptor I (mGluR1) antagonism impairs cocaine-induced conditioned place preference via inhibition of protein synthesis
- PMID: 23348064
- PMCID: PMC3656374
- DOI: 10.1038/npp.2013.29
Metabotropic glutamate receptor I (mGluR1) antagonism impairs cocaine-induced conditioned place preference via inhibition of protein synthesis
Abstract
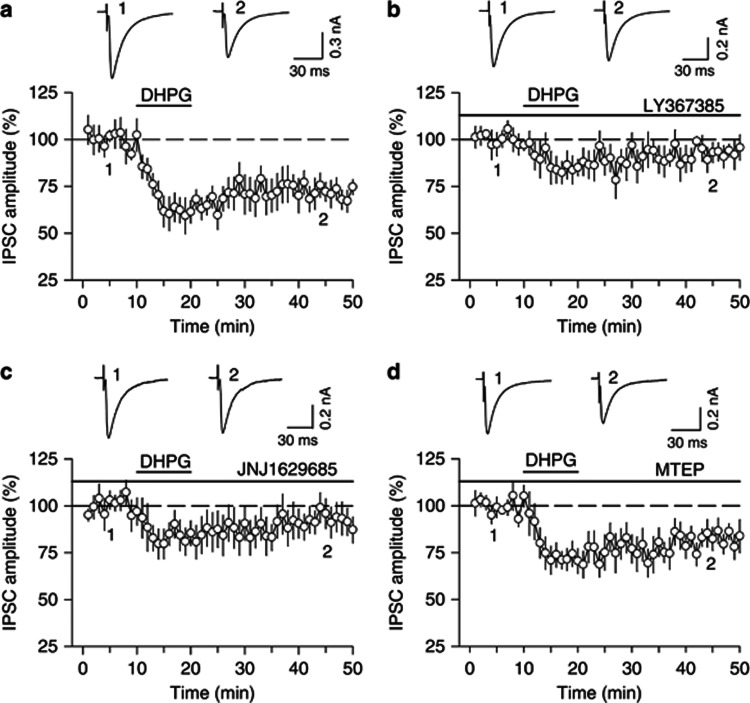
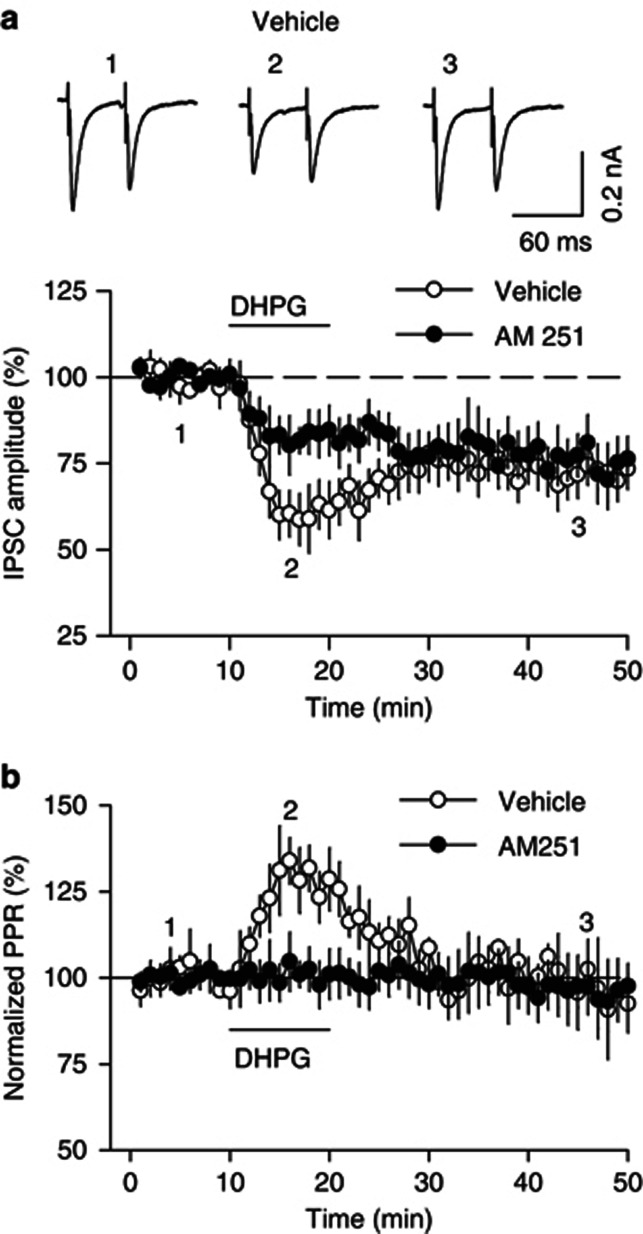
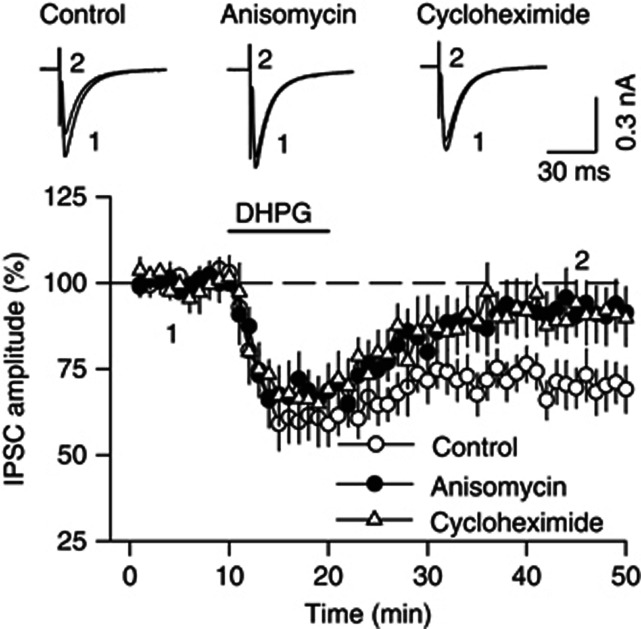
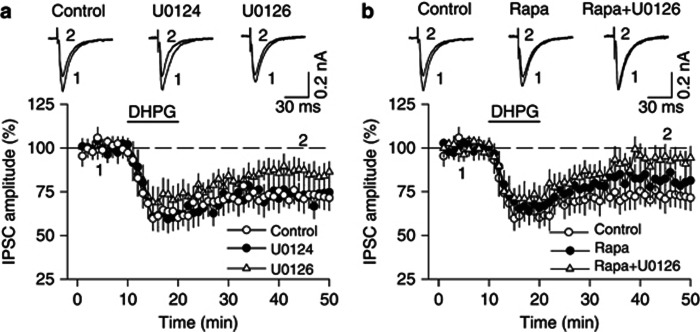
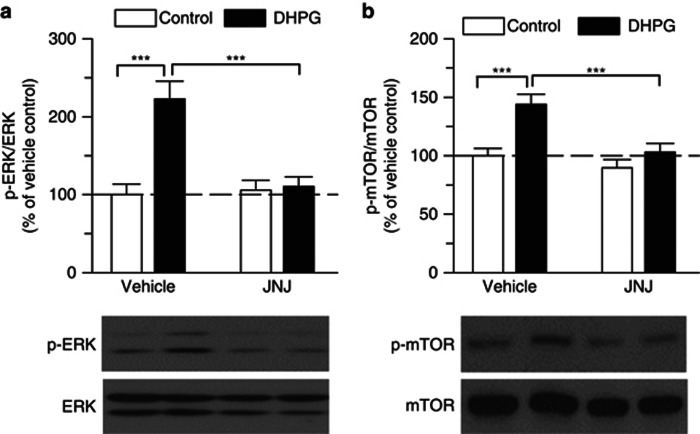
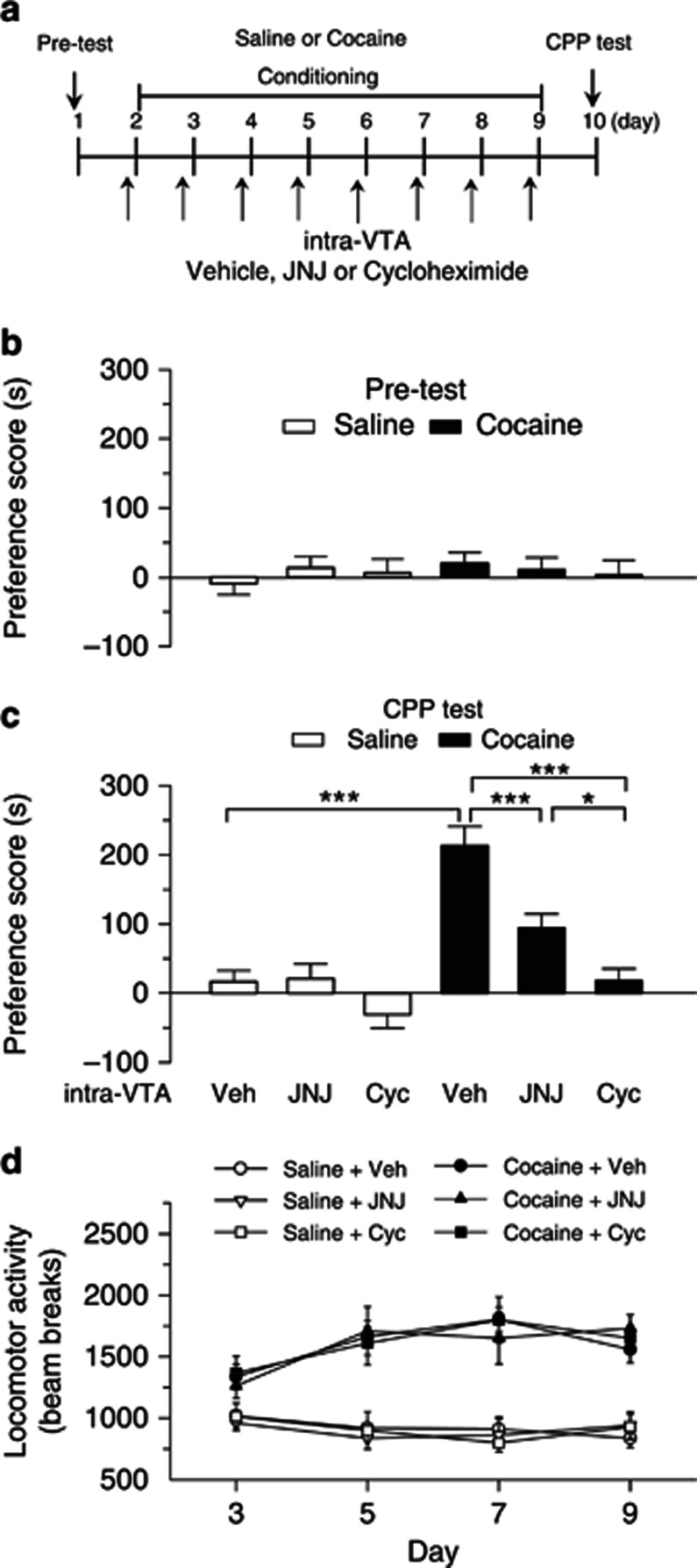
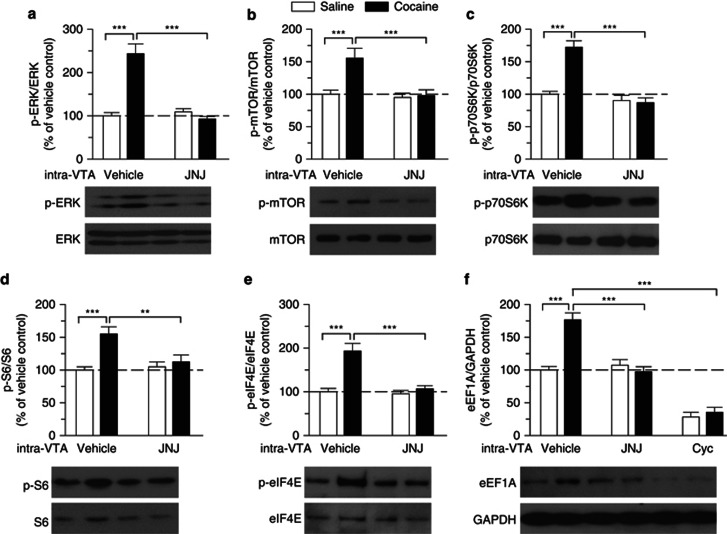
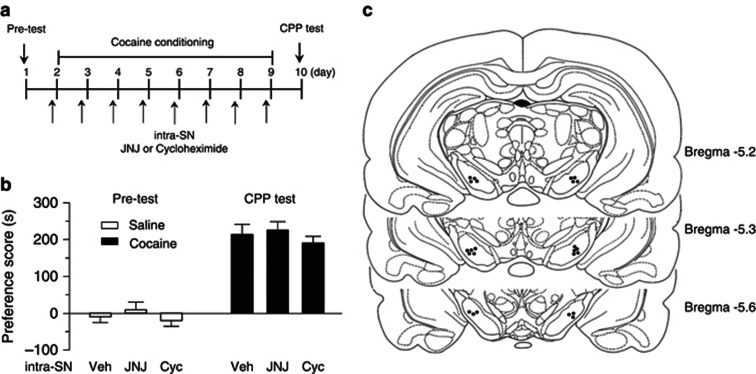
Antagonism of group I metabotropic glutamate receptors (mGluR1 and mGluR5) reduces behavioral effects of drugs of abuse, including cocaine. However, the underlying mechanisms remain poorly understood. Activation of mGluR5 increases protein synthesis at synapses. Although mGluR5-induced excessive protein synthesis has been implicated in the pathology of fragile X syndrome, it remains unknown whether group I mGluR-mediated protein synthesis is involved in any behavioral effects of drugs of abuse. We report that group I mGluR agonist DHPG induced more pronounced initial depression of inhibitory postsynaptic currents (IPSCs) followed by modest long-term depression (I-LTD) in dopamine neurons of rat ventral tegmental area (VTA) through the activation of mGluR1. The early component of DHPG-induced depression of IPSCs was mediated by the cannabinoid CB1 receptors, while DHPG-induced I-LTD was dependent on protein synthesis. Western blotting analysis indicates that mGluR1 was coupled to extracellular signal-regulated kinase (ERK) and mammalian target of rapamycin (mTOR) signaling pathways to increase translation. We also show that cocaine conditioning activated translation machinery in the VTA via an mGluR1-dependent mechanism. Furthermore, intra-VTA microinjections of mGluR1 antagonist JNJ16259685 and protein synthesis inhibitor cycloheximide significantly attenuated or blocked the acquisition of cocaine-induced conditioned place preference (CPP) and activation of translation elongation factors. Taken together, these results suggest that mGluR1 antagonism inhibits de novo protein synthesis; this effect may block the formation of cocaine-cue associations and thus provide a mechanism for the reduction in CPP to cocaine.
Figures
References
-
- Bellone C, Luscher C. mGluRs induce a long-term depression in the ventral tegmental area that involves a switch of the subunit composition of AMPA receptors. Eur J Neurosci. 2005;21:1280–1288. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
