

Caspase-11 protects against bacteria that escape the vacuole
- PMID: 23348507
- PMCID: PMC3697099
- DOI: 10.1126/science.1230751
Caspase-11 protects against bacteria that escape the vacuole
Abstract
Caspases are either apoptotic or inflammatory. Among inflammatory caspases, caspase-1 and -11 trigger pyroptosis, a form of programmed cell death. Whereas both can be detrimental in inflammatory disease, only caspase-1 has an established protective role during infection. Here, we report that caspase-11 is required for innate immunity to cytosolic, but not vacuolar, bacteria. Although Salmonella typhimurium and Legionella pneumophila normally reside in the vacuole, specific mutants (sifA and sdhA, respectively) aberrantly enter the cytosol. These mutants triggered caspase-11, which enhanced clearance of S. typhimurium sifA in vivo. This response did not require NLRP3, NLRC4, or ASC inflammasome pathways. Burkholderia species that naturally invade the cytosol also triggered caspase-11, which protected mice from lethal challenge with B. thailandensis and B. pseudomallei. Thus, caspase-11 is critical for surviving exposure to ubiquitous environmental pathogens.
Figures
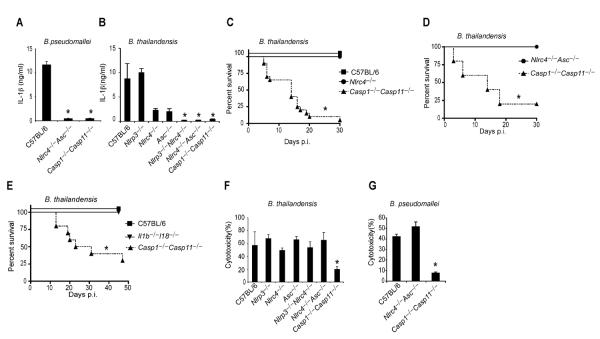
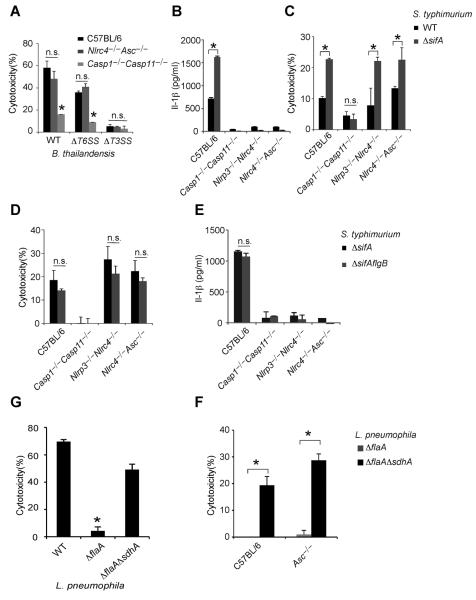
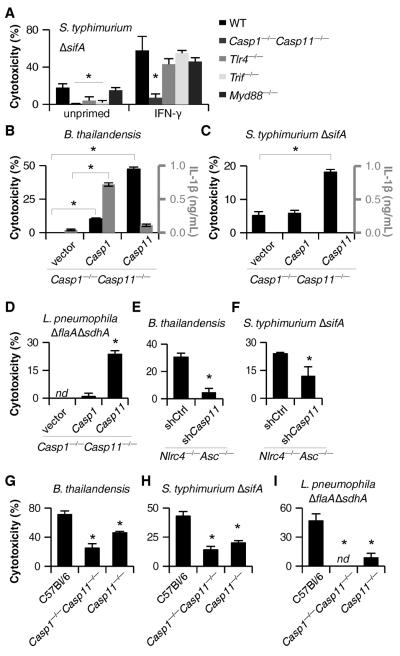
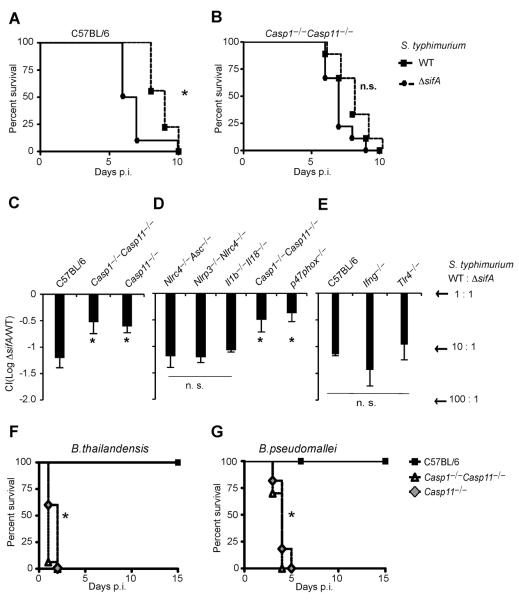
Comment in
-
Innate immunity: Caspase 11 hunts down cytosolic bacteria.Nat Rev Immunol. 2013 Mar;13(3):154-5. doi: 10.1038/nri3408. Epub 2013 Feb 15. Nat Rev Immunol. 2013. PMID: 23411795 No abstract available.
-
Immunology. Bacterial escape artists set afire.Science. 2013 Feb 22;339(6122):912-3. doi: 10.1126/science.1235639. Science. 2013. PMID: 23430642 No abstract available.
-
Caspase-11: the noncanonical guardian of cytosolic sanctity.Cell Host Microbe. 2013 Mar 13;13(3):243-5. doi: 10.1016/j.chom.2013.02.011. Cell Host Microbe. 2013. PMID: 23498948
References
-
- Zhao Y, et al. The NLRC4 inflammasome receptors for bacterial flagellin and type III secretion apparatus. Nature. 2011;477:596–600. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U54 AI057141/AI/NIAID NIH HHS/United States
- AI065359/AI/NIAID NIH HHS/United States
- R37 AI075039/AI/NIAID NIH HHS/United States
- P30 CA016086/CA/NCI NIH HHS/United States
- AI057141/AI/NIAID NIH HHS/United States
- U54 AI065359/AI/NIAID NIH HHS/United States
- AI063302/AI/NIAID NIH HHS/United States
- P01 AI063302/AI/NIAID NIH HHS/United States
- R01 AI097518/AI/NIAID NIH HHS/United States
- R01 AI080749/AI/NIAID NIH HHS/United States
- AI080749/AI/NIAID NIH HHS/United States
- R01 AI075039/AI/NIAID NIH HHS/United States
- AI097518/AI/NIAID NIH HHS/United States
- U19 AI100627/AI/NIAID NIH HHS/United States
- AI075039/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
