

ATP and ADP actin states
- PMID: 23348672
- PMCID: PMC3670783
- DOI: 10.1002/bip.22155
ATP and ADP actin states
Abstract
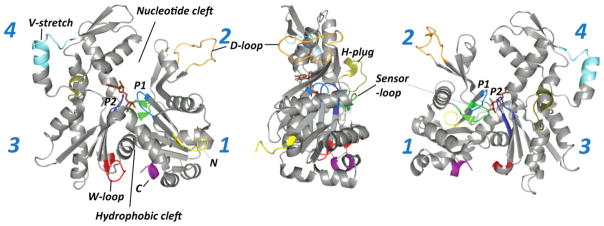
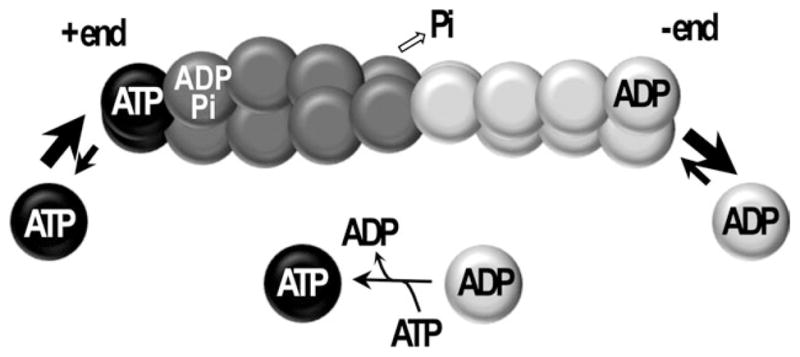
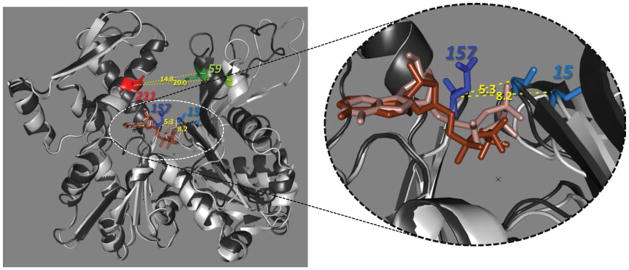
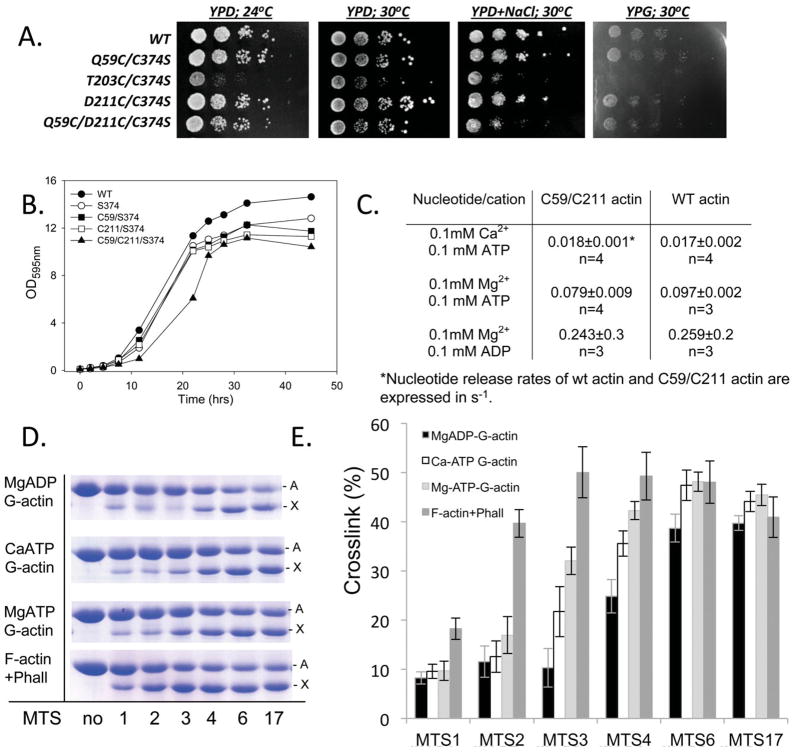
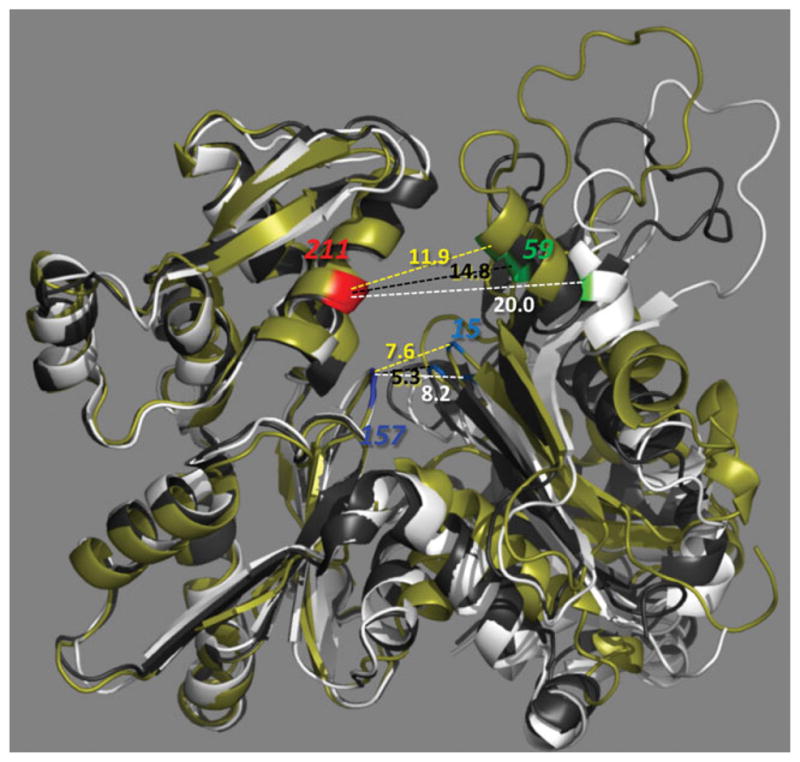
This minireview is dedicated to the memory of Henryk Eisenberg and honors his major contributions to many areas of biophysics and to the analysis of macromolecular states and interactions in particular. This work reviews the ATP and ADP states of a ubiquitous protein, actins, and considers the present evidence for and against unique, nucleotide-dependent conformations of this protein. The effects of ATP and ADP on specific structural elements of actins, its loops and clefts, as revealed by mutational, crosslinking, spectroscopic, and EPR methods are discussed. It is concluded that the existing evidence points to dynamic equilibria of these structural elements among various conformational states in both ATP- and ADP-actins, with the nucleotides impacting the equilibria distributions.
Copyright © 2012 Wiley Periodicals, Inc.
Figures
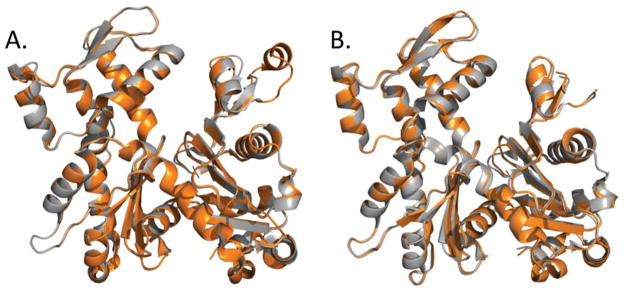
Similar articles
-
Replacement of ATP with ADP affects the dynamic and conformational properties of actin monomer.Biochemistry. 1999 Sep 28;38(39):12885-92. doi: 10.1021/bi990748y. Biochemistry. 1999. PMID: 10504259
-
Conformational and dynamic differences between actin filaments polymerized from ATP- or ADP-actin monomers.J Biol Chem. 2000 Dec 29;275(52):41143-9. doi: 10.1074/jbc.M004146200. J Biol Chem. 2000. PMID: 11005806
-
Partial specific volume and adiabatic compressibility of G-actin depend on the bound nucleotide.J Biochem. 2003 May;133(5):687-91. doi: 10.1093/jb/mvg088. J Biochem. 2003. PMID: 12801922
-
Nucleotide-dependence of G-actin conformation from multiple molecular dynamics simulations and observation of a putatively polymerization-competent superclosed state.Proteins. 2009 Aug 1;76(2):353-64. doi: 10.1002/prot.22350. Proteins. 2009. PMID: 19156817
-
Nucleotide-dependent conformational states of actin.Proc Natl Acad Sci U S A. 2009 Aug 4;106(31):12723-8. doi: 10.1073/pnas.0902092106. Epub 2009 Jul 20. Proc Natl Acad Sci U S A. 2009. PMID: 19620726 Free PMC article.
Cited by
-
Structural insights into actin filament recognition by commonly used cellular actin markers.EMBO J. 2020 Jul 15;39(14):e104006. doi: 10.15252/embj.2019104006. Epub 2020 Jun 22. EMBO J. 2020. PMID: 32567727 Free PMC article.
-
Oligomerization Affects the Ability of Human Cyclase-Associated Proteins 1 and 2 to Promote Actin Severing by Cofilins.Int J Mol Sci. 2019 Nov 12;20(22):5647. doi: 10.3390/ijms20225647. Int J Mol Sci. 2019. PMID: 31718088 Free PMC article.
-
Effect of the Rho-Kinase/ROCK Signaling Pathway on Cytoskeleton Components.Genes (Basel). 2023 Jan 20;14(2):272. doi: 10.3390/genes14020272. Genes (Basel). 2023. PMID: 36833199 Free PMC article. Review.
-
Actin age orchestrates myosin-5 and myosin-6 run lengths.Curr Biol. 2015 Aug 3;25(15):2057-62. doi: 10.1016/j.cub.2015.06.033. Epub 2015 Jul 16. Curr Biol. 2015. PMID: 26190073 Free PMC article.
-
Nucleotide regulation of the structure and dynamics of G-actin.Biophys J. 2014 Apr 15;106(8):1710-20. doi: 10.1016/j.bpj.2014.03.012. Biophys J. 2014. PMID: 24739170 Free PMC article.
References
-
- Straub FB. Studies Inst Med Chem Univ Szeged. 1942;2:3–15.
-
- Straub FB. Studies Inst Med Chem Univ Szeged. 1943;3:23–37.
-
- Szent-György A. Acta Physiol Scand. 1945;9:25.
-
- Pollard TD, Shelton E, Weihing RR, Korn ED. J Mol Biol. 1970;50:91–97. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
