

Store-independent Orai1/3 channels activated by intracrine leukotriene C4: role in neointimal hyperplasia
- PMID: 23349245
- PMCID: PMC3658112
- DOI: 10.1161/CIRCRESAHA.111.300220
Store-independent Orai1/3 channels activated by intracrine leukotriene C4: role in neointimal hyperplasia
Abstract
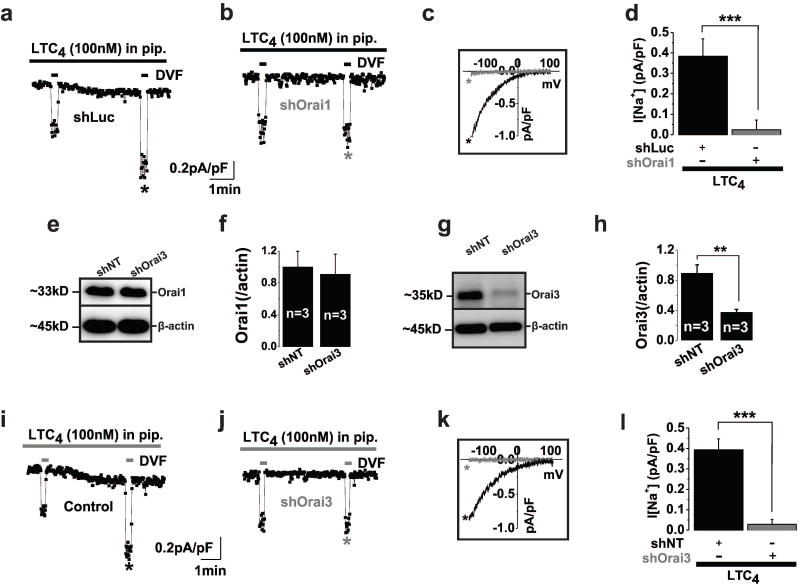
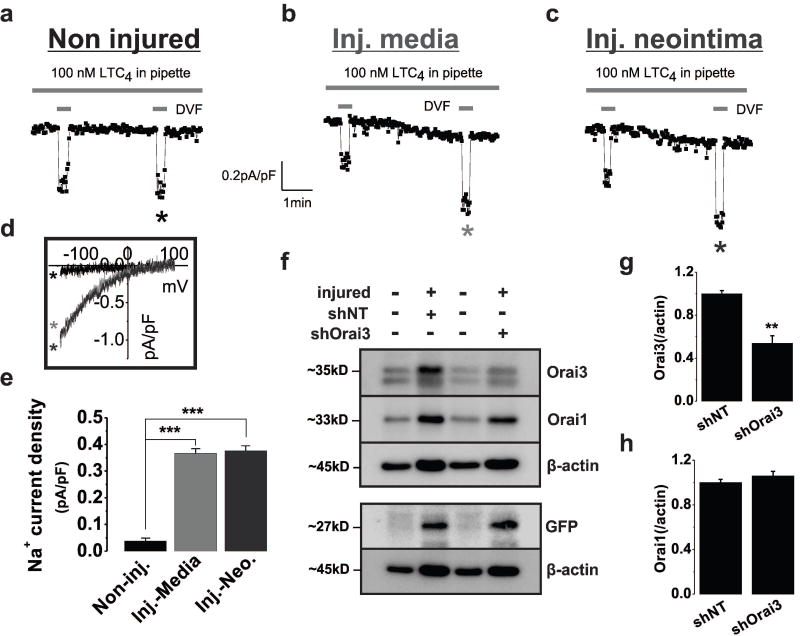
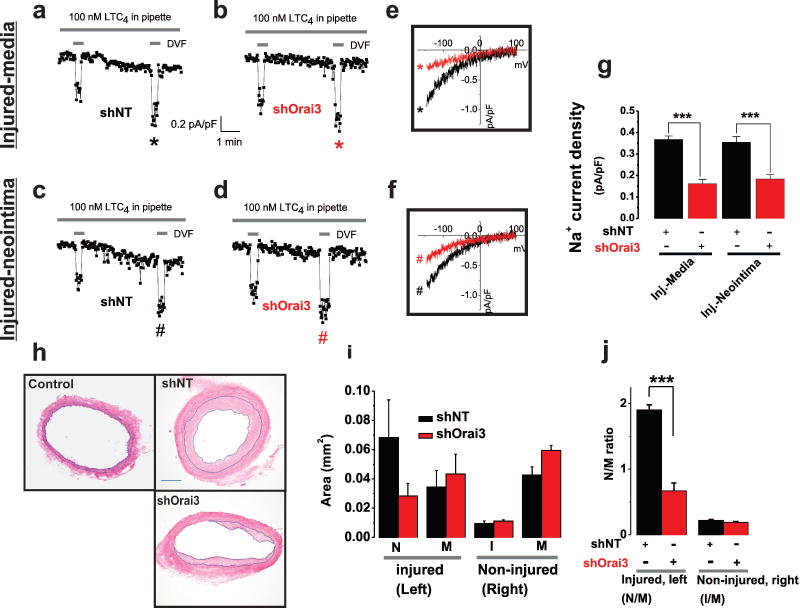
Rationale: Through largely unknown mechanisms, Ca(2+) signaling plays important roles in vascular smooth muscle cell (VSMC) remodeling. Orai1-encoded store-operated Ca(2+) entry has recently emerged as an important player in VSMC remodeling. However, the role of the exclusively mammalian Orai3 protein in native VSMC Ca(2+) entry pathways, its upregulation during VSMC remodeling, and its contribution to neointima formation remain unknown.
Objective: The goal of this study was to determine the agonist-evoked Ca(2+) entry pathway contributed by Orai3; Orai3 potential upregulation and role during neointima formation after balloon injury of rat carotid arteries.
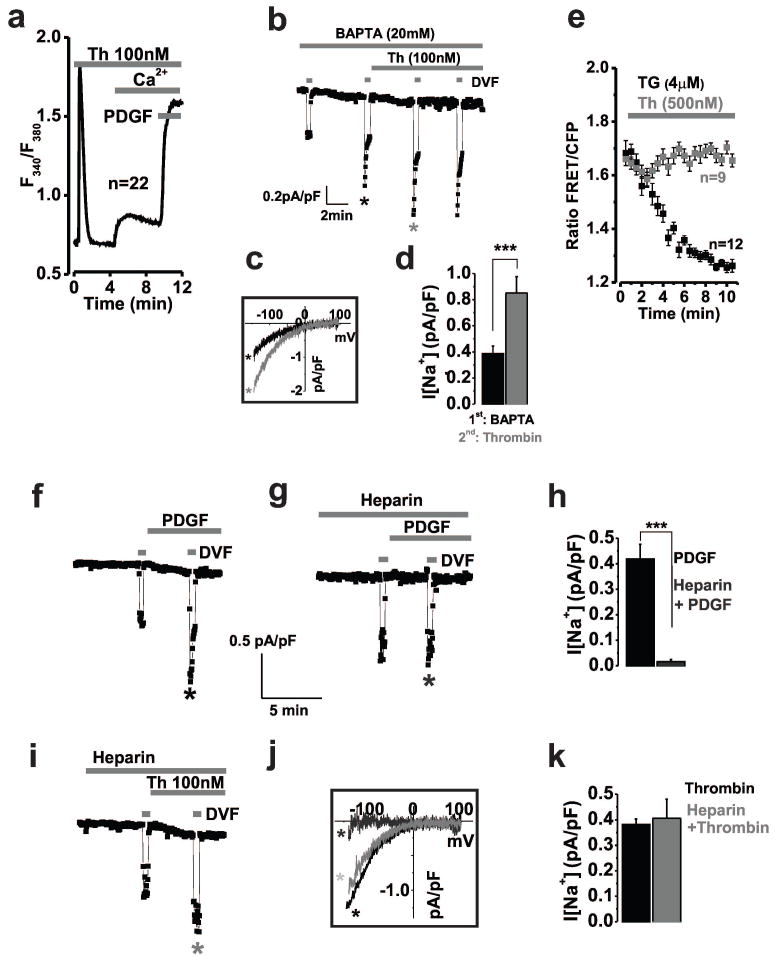
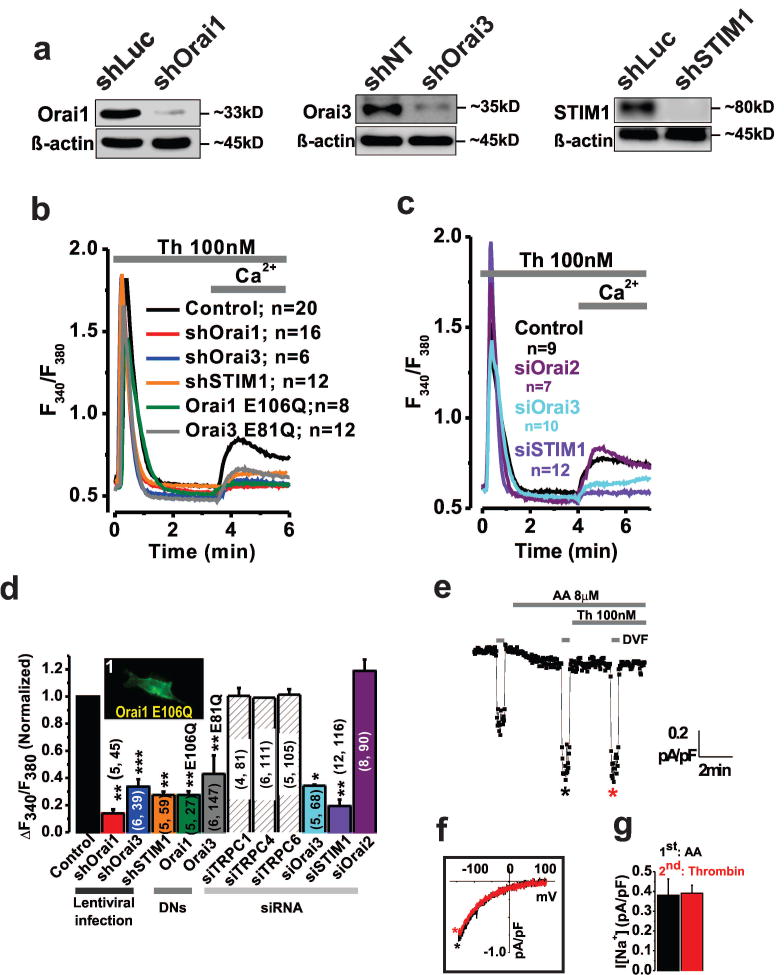
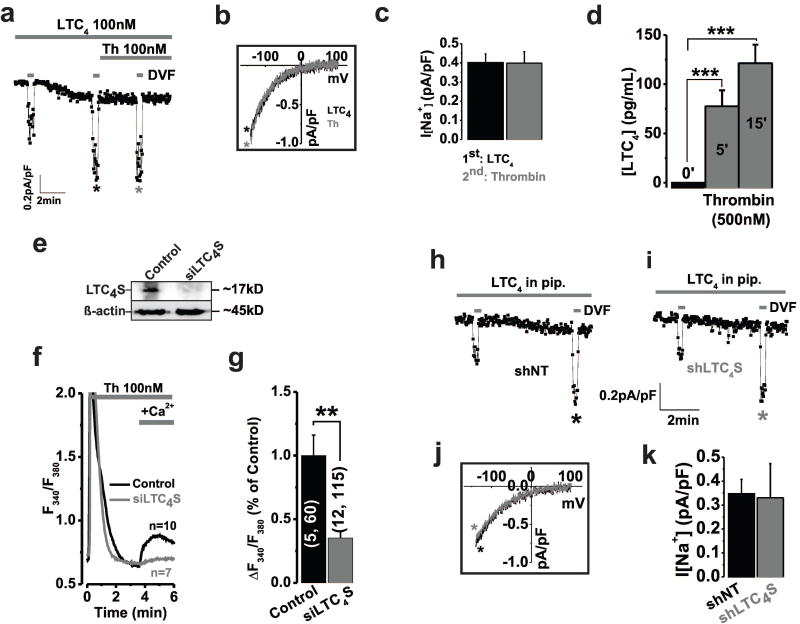
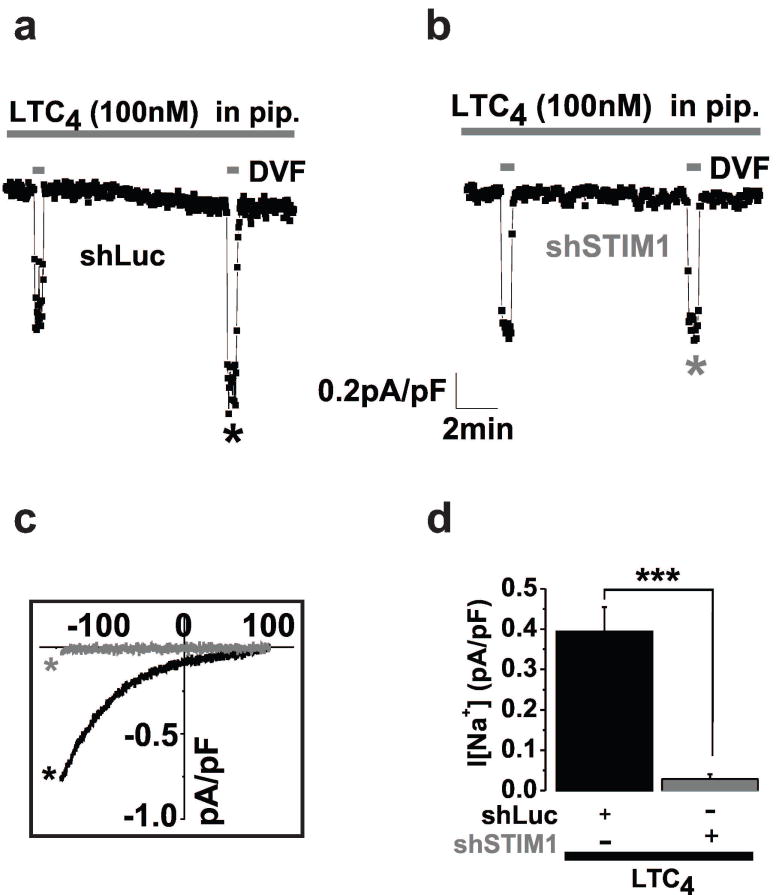
Methods and results: Ca(2+) imaging and patch-clamp recordings showed that although the platelet-derived growth factor activates the canonical Ca(2+) release-activated Ca(2+) channels via store depletion in VSMC, the pathophysiological agonist thrombin activates a distinct Ca(2+)-selective channel contributed by Orai1, Orai3, and stromal interacting molecule1 in the same cells. Unexpectedly, Ca(2+) store depletion is not required for activation of Orai1/3 channel by thrombin. Rather, the signal for Orai1/3 channel activation is cytosolic leukotrieneC4 produced downstream thrombin receptor stimulation through the catalytic activity of leukotrieneC4 synthase. Importantly, Orai3 is upregulated in an animal model of VSMC neointimal remodeling, and in vivo Orai3 knockdown inhibits neointima formation.
Conclusions: These results demonstrate that distinct native Ca(2+)-selective Orai channels are activated by different agonists/pathways and uncover a mechanism whereby leukotrieneC4 acts through hitherto unknown intracrine mode to elicit store-independent Ca(2+) signaling that promotes vascular occlusive disease. Orai3 and Orai3-containing channels provide novel targets for control of VSMC remodeling during vascular injury or disease.
Figures
Comment in
-
The many faces of Orai.Circ Res. 2013 Mar 29;112(7):983-4. doi: 10.1161/CIRCRESAHA.113.301052. Circ Res. 2013. PMID: 23538273 No abstract available.
References
-
- Berridge MJ. Calcium microdomains: Organization and function. Cell calcium. 2006;40:405–412. - PubMed
-
- Clapham DE. Calcium signaling. Cell. 2007;131:1047–1058. - PubMed
-
- Putney JW., Jr A model for receptor-regulated calcium entry. Cell calcium. 1986;7:1–12. - PubMed
-
- Bird GS, Aziz O, Lievremont JP, Wedel BJ, Trebak M, Vazquez G, Putney JW., Jr Mechanisms of phospholipase c-regulated calcium entry. Current molecular medicine. 2004;4:291–301. - PubMed
-
- Hoth M, Penner R. Depletion of intracellular calcium stores activates a calcium current in mast cells. Nature. 1992;355:353–356. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
