

Erythrocyte NADPH oxidase activity modulated by Rac GTPases, PKC, and plasma cytokines contributes to oxidative stress in sickle cell disease
- PMID: 23349388
- PMCID: PMC3596970
- DOI: 10.1182/blood-2012-07-441188
Erythrocyte NADPH oxidase activity modulated by Rac GTPases, PKC, and plasma cytokines contributes to oxidative stress in sickle cell disease
Erratum in
- Blood. 2014 Mar 20;123(12):1972
Abstract
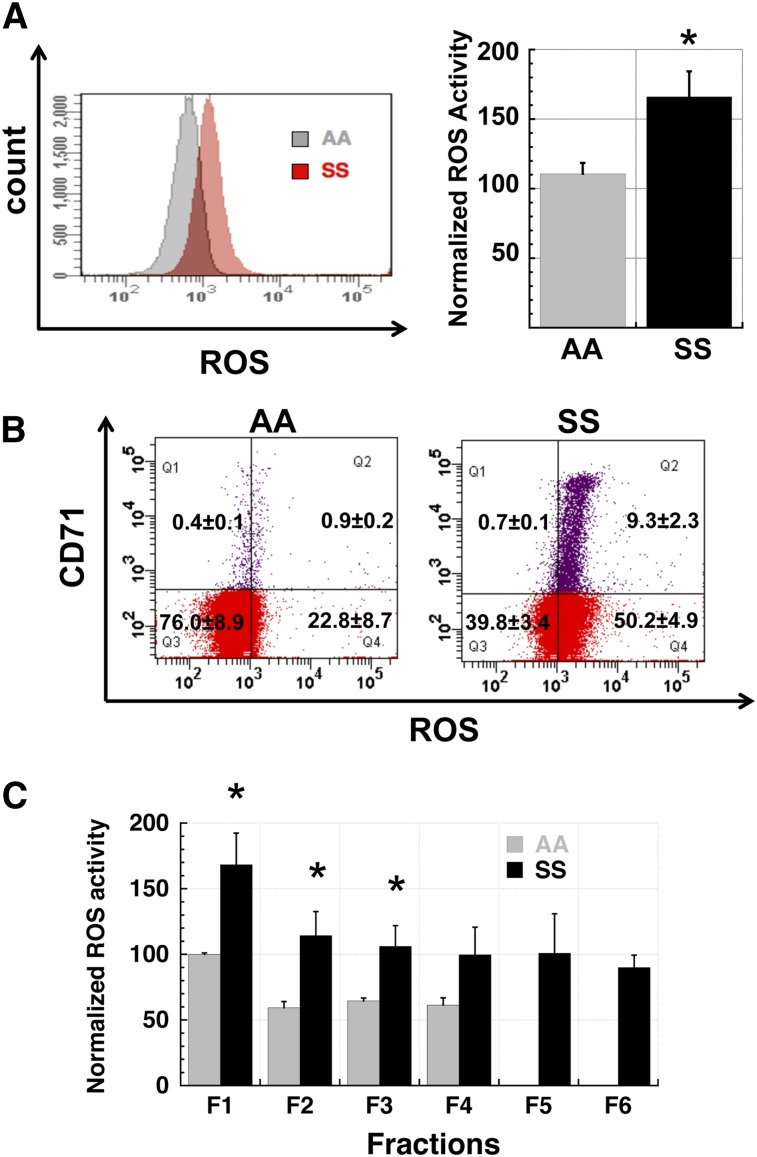
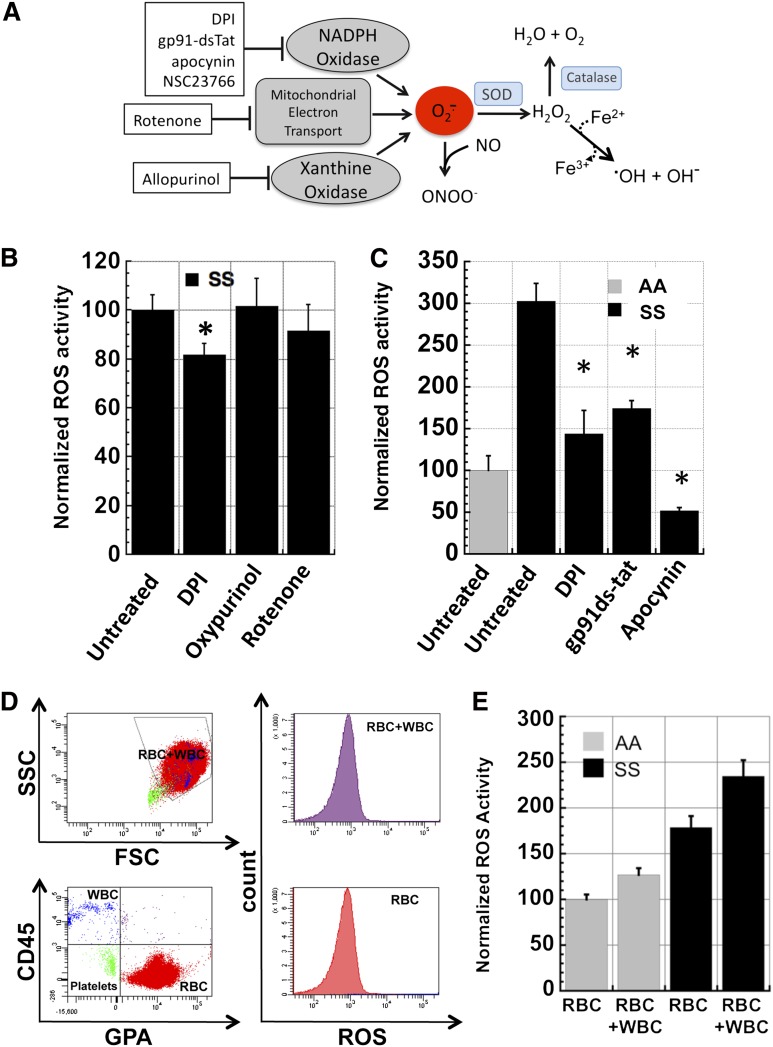
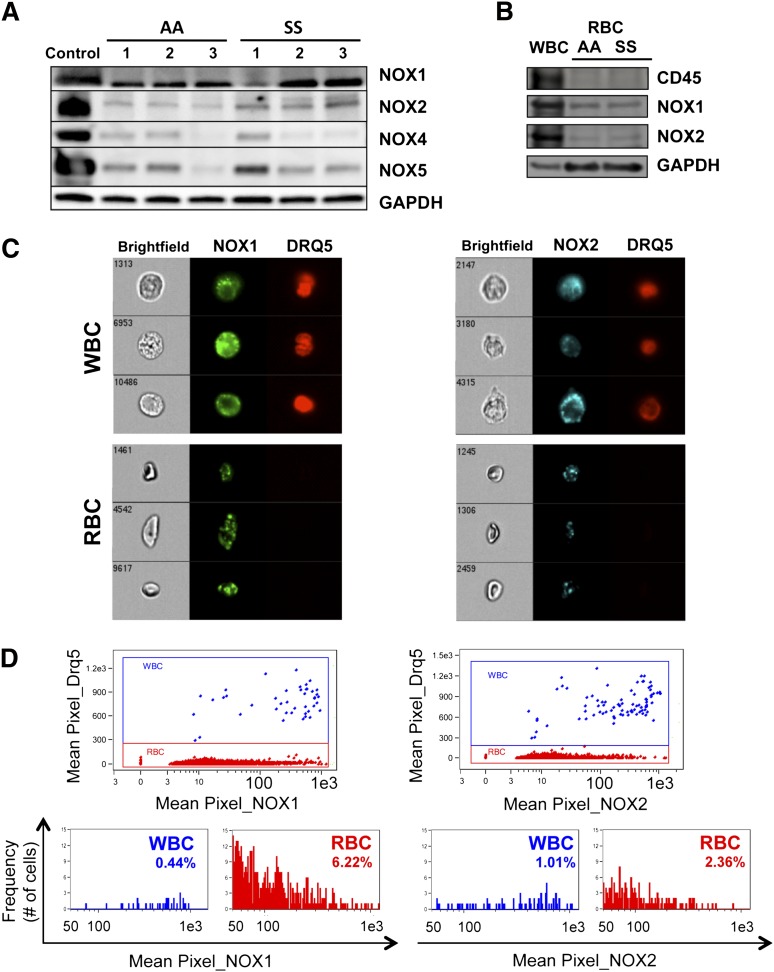
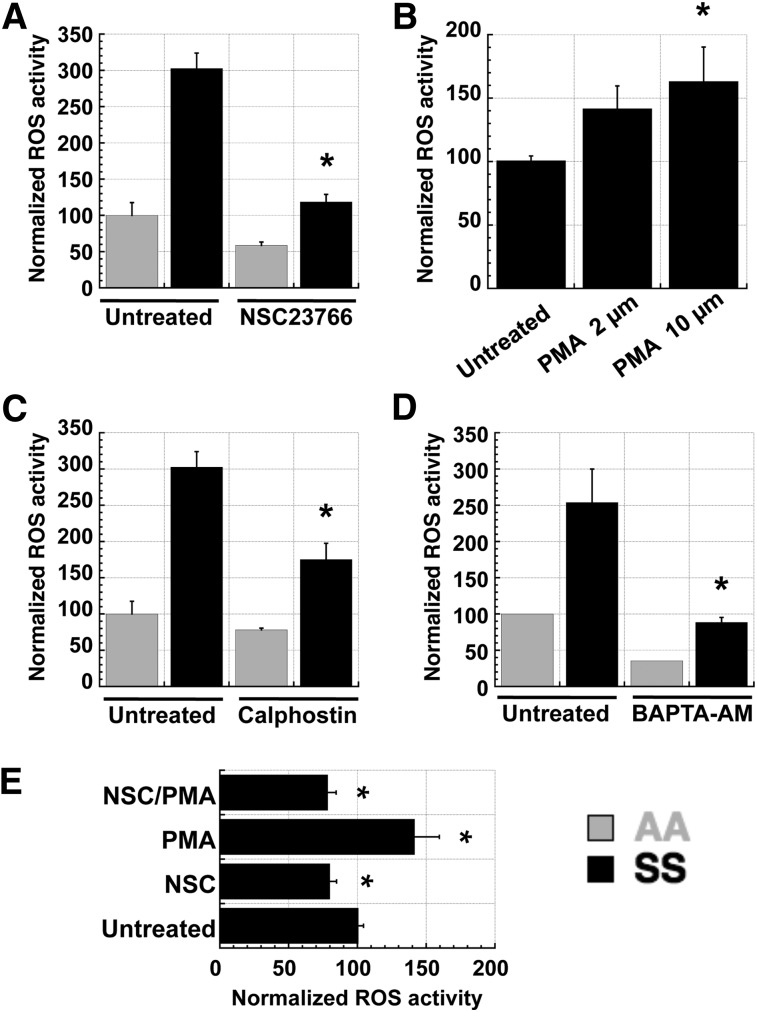
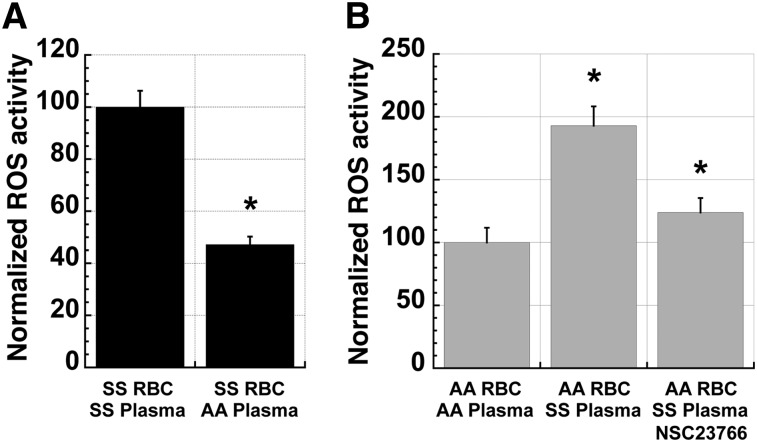
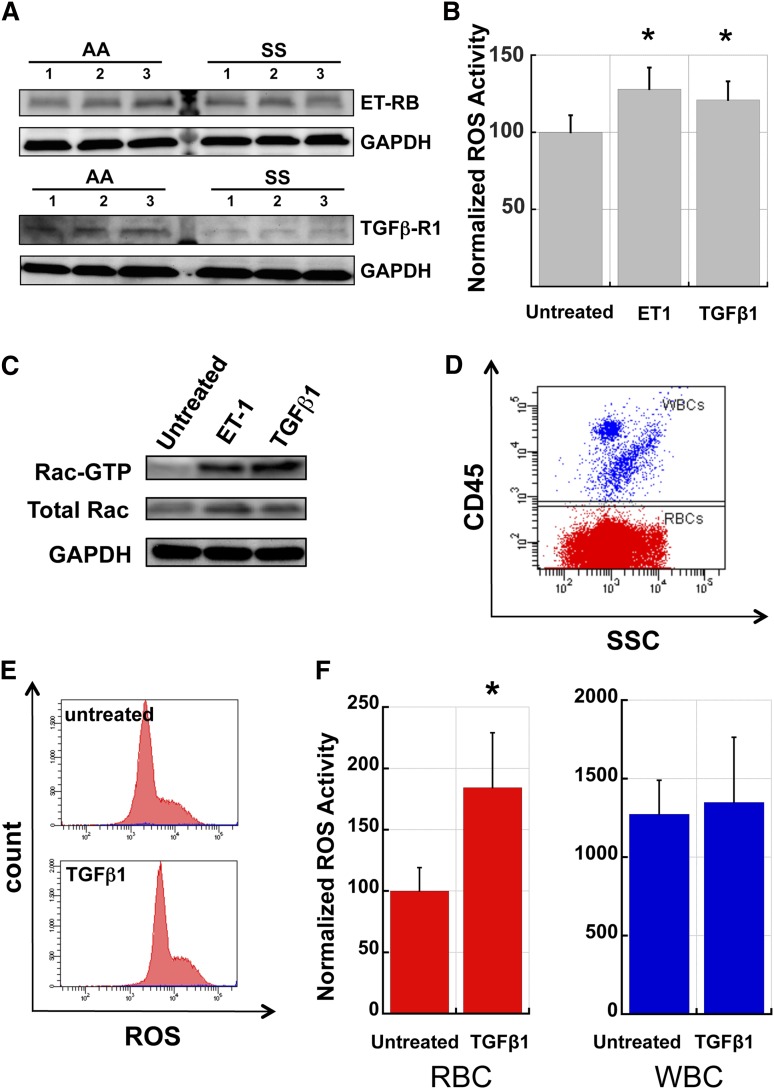
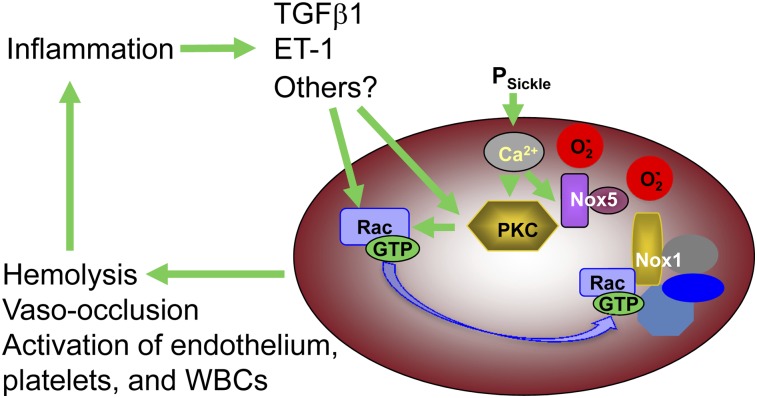
Chronic inflammation has emerged as an important pathogenic mechanism in sickle cell disease (SCD). One component of this inflammatory response is oxidant stress mediated by reactive oxygen species (ROS) generated by leukocytes, endothelial cells, plasma enzymes, and sickle red blood cells (RBC). Sickle RBC ROS generation has been attributed to sickle hemoglobin auto-oxidation and Fenton chemistry reactions catalyzed by denatured heme moieties bound to the RBC membrane. In this study, we demonstrate that a significant part of ROS production in sickle cells is mediated enzymatically by NADPH oxidase, which is regulated by protein kinase C, Rac GTPase, and intracellular Ca(2+) signaling within the sickle RBC. Moreover, plasma from patients with SCD and isolated cytokines, such as transforming growth factor β1 and endothelin-1, enhance RBC NADPH oxidase activity and increase ROS generation. ROS-mediated damage to RBC membrane components is known to contribute to erythrocyte rigidity and fragility in SCD. Erythrocyte ROS generation, hemolysis, vaso-occlusion, and the inflammatory response to tissue damage may therefore act in a positive-feedback loop to drive the pathophysiology of sickle cell disease. These findings suggest a novel pathogenic mechanism in SCD and may offer new therapeutic targets to counteract inflammation and RBC rigidity and fragility in SCD.
Figures
References
-
- Nur E, Biemond BJ, Otten HM, et al. CURAMA Study Group. Oxidative stress in sickle cell disease; pathophysiology and potential implications for disease management. Am J Hematol. 2011;86(6):484–489. - PubMed
-
- Ray D, Deshmukh P, Goswami K, et al. Antioxidant vitamin levels in sickle cell disorders. Natl Med J India. 2007;20(1):11–13. - PubMed
-
- Ren H, Ghebremeskel K, Okpala I, et al. Patients with sickle cell disease have reduced blood antioxidant protection. Int J Vitam Nutr Res. 2008;78(3):139–147. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01HL116352/HL/NHLBI NIH HHS/United States
- K08HL088126/HL/NHLBI NIH HHS/United States
- DK26263/DK/NIDDK NIH HHS/United States
- U54 HL070871/HL/NHLBI NIH HHS/United States
- R01 DK026263/DK/NIDDK NIH HHS/United States
- K08 HL088126/HL/NHLBI NIH HHS/United States
- R37 DK026263/DK/NIDDK NIH HHS/United States
- P30 DK090971/DK/NIDDK NIH HHS/United States
- U01 HL117709/HL/NHLBI NIH HHS/United States
- R01 HL116352/HL/NHLBI NIH HHS/United States
- P30DK090971/DK/NIDDK NIH HHS/United States
- U54HL070871/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
