

LKB1 regulates lipid oxidation during exercise independently of AMPK
- PMID: 23349504
- PMCID: PMC3636614
- DOI: 10.2337/db12-1160
LKB1 regulates lipid oxidation during exercise independently of AMPK
Abstract
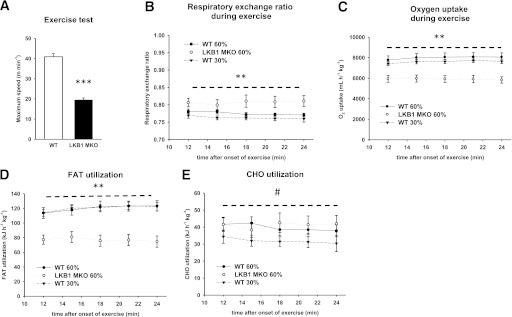
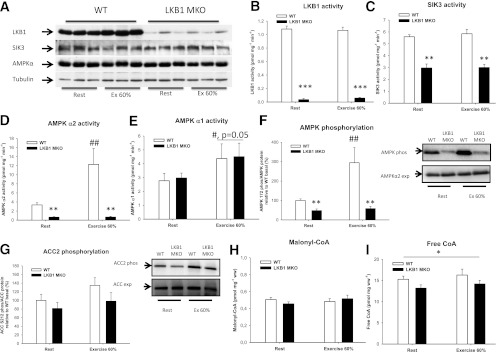
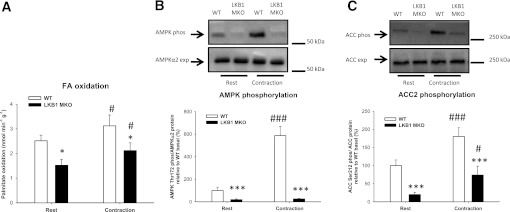
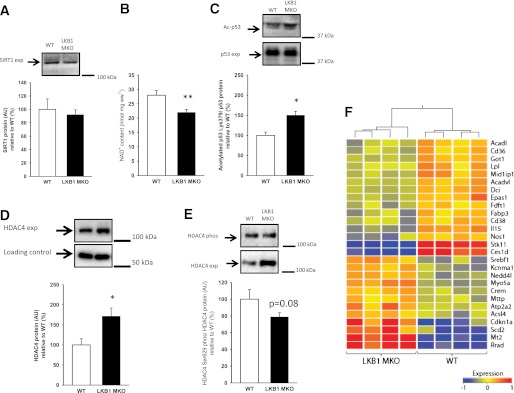
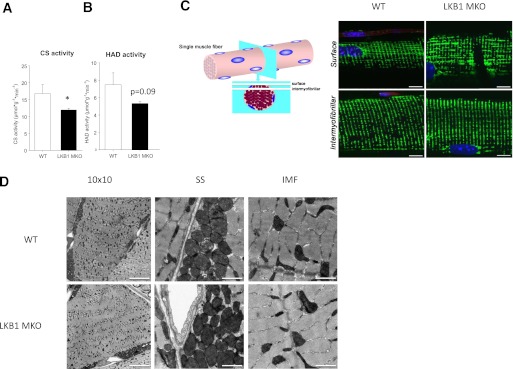
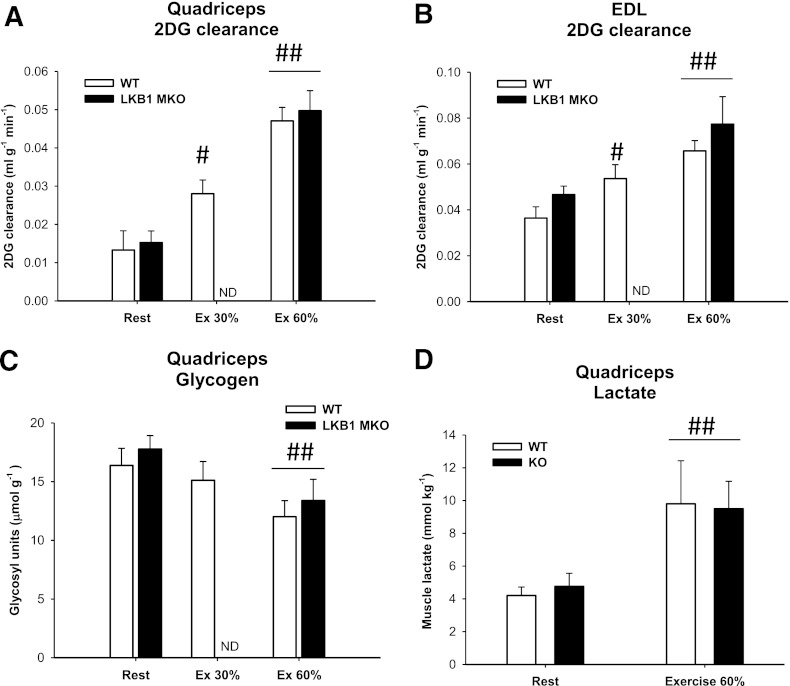
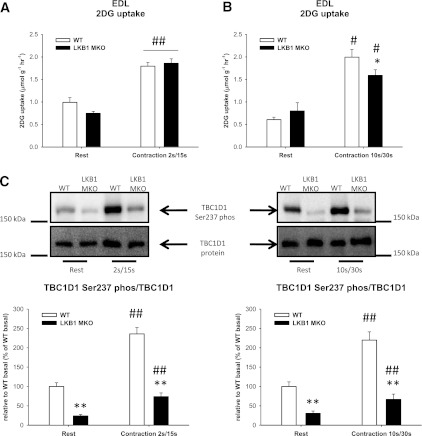
Lipid metabolism is important for health and insulin action, yet the fundamental process of regulating lipid metabolism during muscle contraction is incompletely understood. Here, we show that liver kinase B1 (LKB1) muscle-specific knockout (LKB1 MKO) mice display decreased fatty acid (FA) oxidation during treadmill exercise. LKB1 MKO mice also show decreased muscle SIK3 activity, increased histone deacetylase 4 expression, decreased NAD⁺ concentration and SIRT1 activity, and decreased expression of genes involved in FA oxidation. In AMP-activated protein kinase (AMPK)α2 KO mice, substrate use was similar to that in WT mice, which excluded that decreased FA oxidation in LKB1 MKO mice was due to decreased AMPKα2 activity. Additionally, LKB1 MKO muscle demonstrated decreased FA oxidation in vitro. A markedly decreased phosphorylation of TBC1D1, a proposed regulator of FA transport, and a low CoA content could contribute to the low FA oxidation in LKB1 MKO. LKB1 deficiency did not reduce muscle glucose uptake or oxidation during exercise in vivo, excluding a general impairment of substrate use during exercise in LKB1 MKO mice. Our findings demonstrate that LKB1 is a novel molecular regulator of major importance for FA oxidation but not glucose uptake in muscle during exercise.
Figures
Similar articles
-
LKB1 and the regulation of malonyl-CoA and fatty acid oxidation in muscle.Am J Physiol Endocrinol Metab. 2007 Dec;293(6):E1572-9. doi: 10.1152/ajpendo.00371.2007. Epub 2007 Oct 9. Am J Physiol Endocrinol Metab. 2007. PMID: 17925454
-
Marked phenotypic differences of endurance performance and exercise-induced oxygen consumption between AMPK and LKB1 deficiency in mouse skeletal muscle: changes occurring in the diaphragm.Am J Physiol Endocrinol Metab. 2013 Jul 15;305(2):E213-29. doi: 10.1152/ajpendo.00114.2013. Epub 2013 May 21. Am J Physiol Endocrinol Metab. 2013. PMID: 23695215
-
LKB1 and AMPK and the regulation of skeletal muscle metabolism.Curr Opin Clin Nutr Metab Care. 2008 May;11(3):227-32. doi: 10.1097/MCO.0b013e3282fb7b76. Curr Opin Clin Nutr Metab Care. 2008. PMID: 18403917 Free PMC article. Review.
-
5'-AMP activated protein kinase α2 controls substrate metabolism during post-exercise recovery via regulation of pyruvate dehydrogenase kinase 4.J Physiol. 2015 Nov 1;593(21):4765-80. doi: 10.1113/JP270821. J Physiol. 2015. PMID: 26359931 Free PMC article.
-
Targeting AMP-activated protein kinase as a novel therapeutic approach for the treatment of metabolic disorders.Diabetes Metab. 2007 Dec;33(6):395-402. doi: 10.1016/j.diabet.2007.10.004. Epub 2007 Nov 7. Diabetes Metab. 2007. PMID: 17997341 Review.
Cited by
-
Activation of AMPKα2 Is Not Required for Mitochondrial FAT/CD36 Accumulation during Exercise.PLoS One. 2015 May 12;10(5):e0126122. doi: 10.1371/journal.pone.0126122. eCollection 2015. PLoS One. 2015. PMID: 25965390 Free PMC article.
-
Investigation of LKB1 Ser431 phosphorylation and Cys433 farnesylation using mouse knockin analysis reveals an unexpected role of prenylation in regulating AMPK activity.Biochem J. 2014 Feb 15;458(1):41-56. doi: 10.1042/BJ20131324. Biochem J. 2014. PMID: 24295069 Free PMC article.
-
Lkb1 controls brown adipose tissue growth and thermogenesis by regulating the intracellular localization of CRTC3.Nat Commun. 2016 Jul 27;7:12205. doi: 10.1038/ncomms12205. Nat Commun. 2016. PMID: 27461402 Free PMC article.
-
Adipocyte-specific DKO of Lkb1 and mTOR protects mice against HFD-induced obesity, but results in insulin resistance.J Lipid Res. 2018 Jun;59(6):974-981. doi: 10.1194/jlr.M081463. Epub 2018 Apr 10. J Lipid Res. 2018. PMID: 29636366 Free PMC article.
-
The tumour suppressor LKB1 regulates myelination through mitochondrial metabolism.Nat Commun. 2014 Sep 26;5:4993. doi: 10.1038/ncomms5993. Nat Commun. 2014. PMID: 25256100 Free PMC article.
References
-
- Richter EA, Mikines KJ, Galbo H, Kiens B. Effect of exercise on insulin action in human skeletal muscle. J Appl Physiol 1989;66:876–885 - PubMed
-
- Merrill GF, Kurth EJ, Hardie DG, Winder WW. AICA riboside increases AMP-activated protein kinase, fatty acid oxidation, and glucose uptake in rat muscle. Am J Physiol 1997;273:E1107–E1112 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
