

The effects of methionine acquisition and synthesis on Streptococcus pneumoniae growth and virulence
- PMID: 23349662
- PMCID: PMC3551916
- DOI: 10.1371/journal.pone.0049638
The effects of methionine acquisition and synthesis on Streptococcus pneumoniae growth and virulence
Abstract
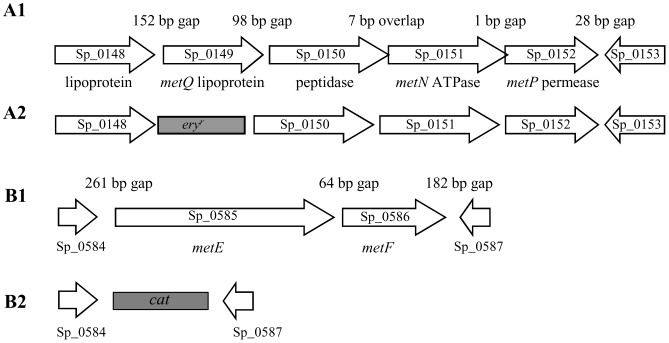
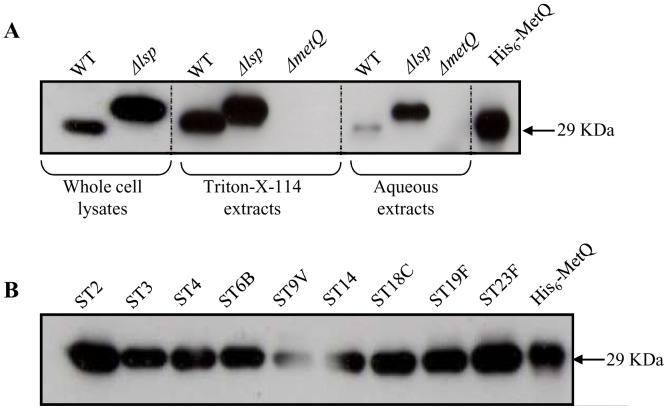
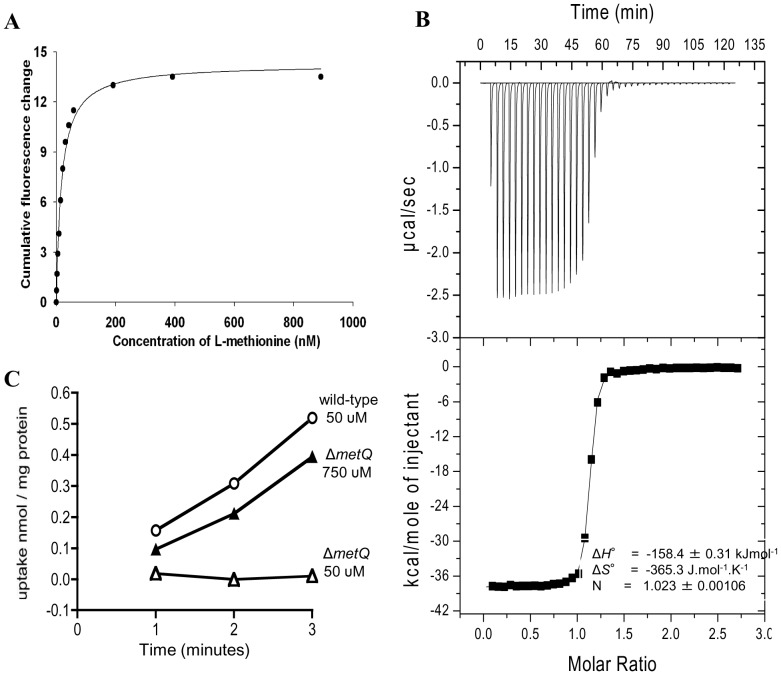
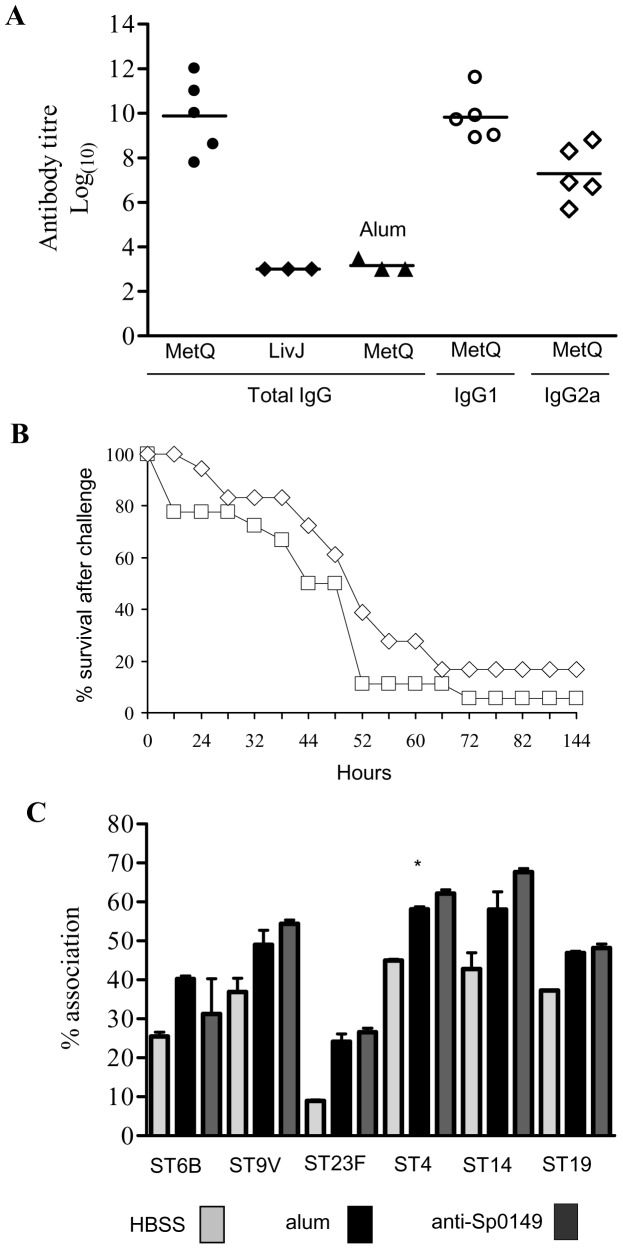
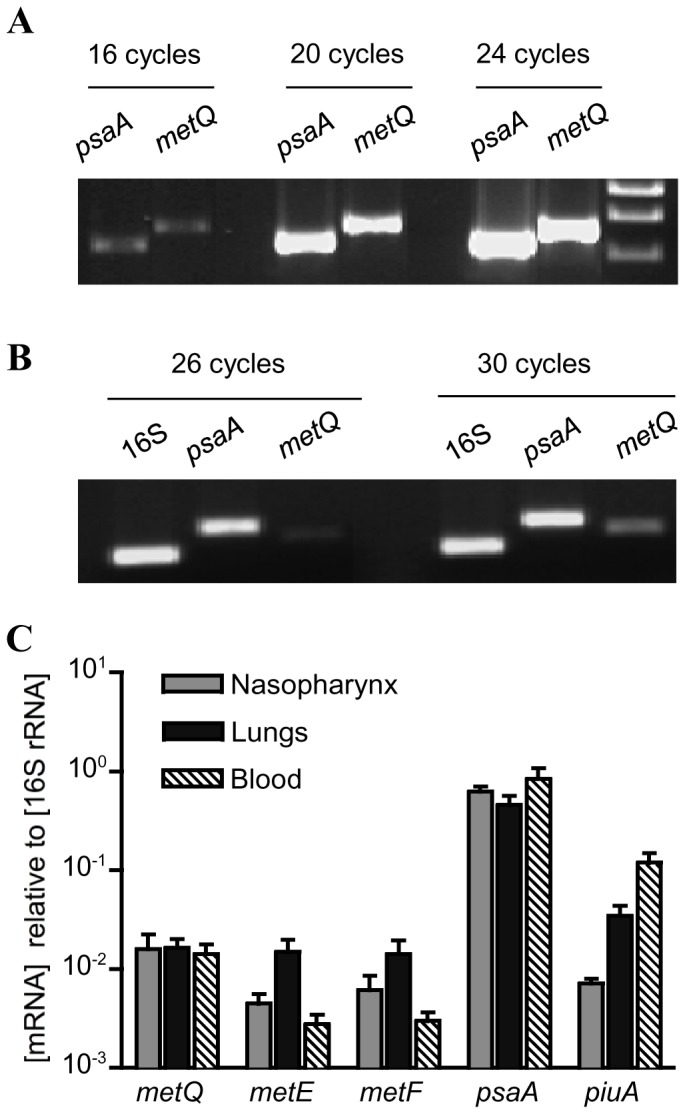
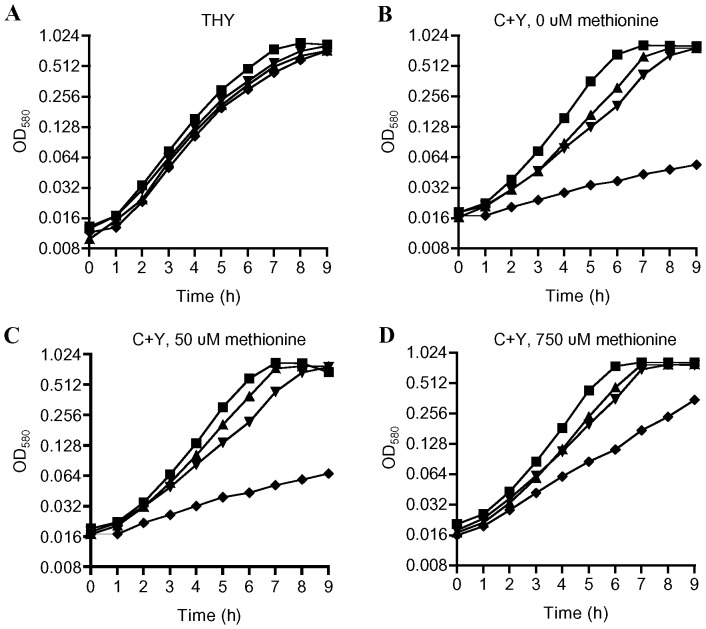
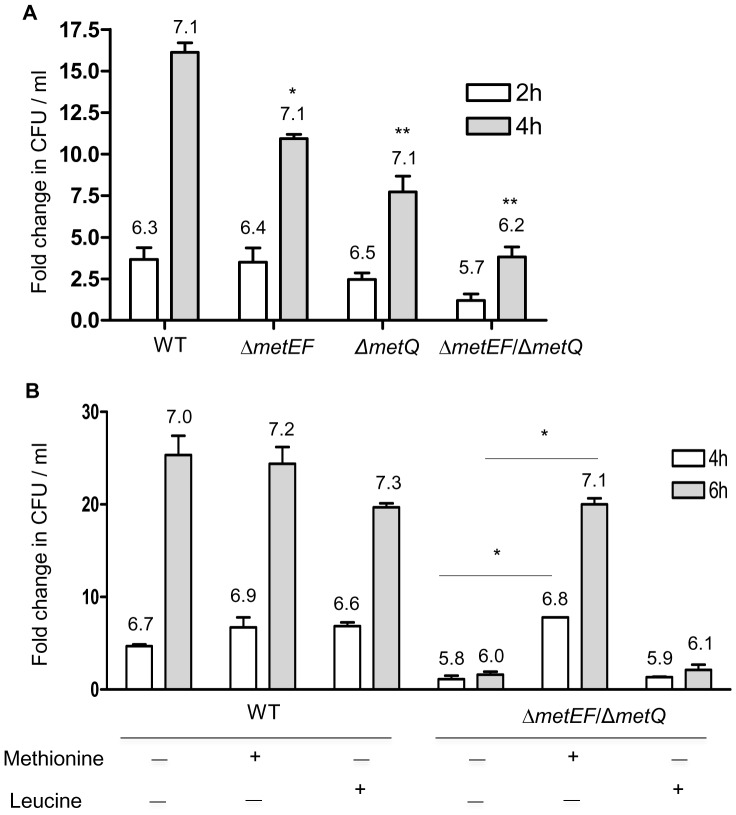
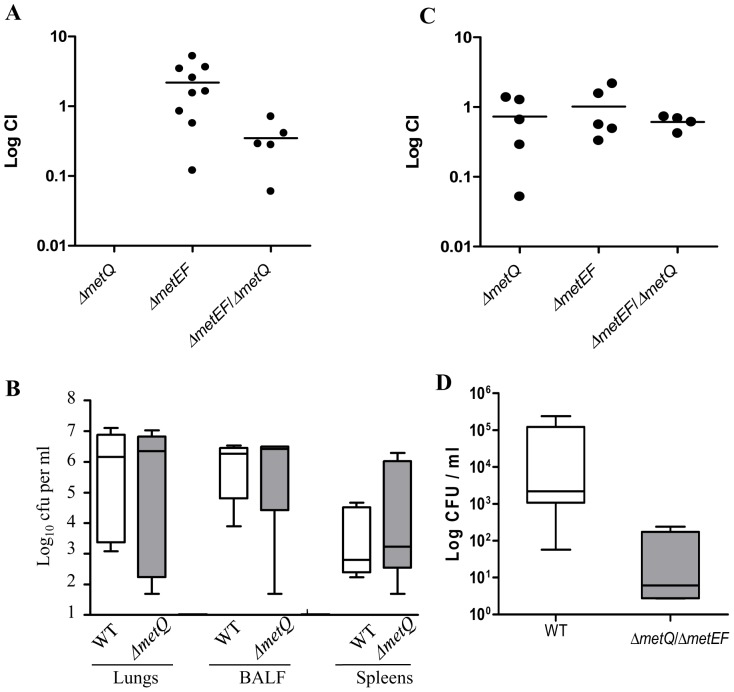
Bacterial pathogens need to acquire nutrients from the host, but for many nutrients their importance during infection remain poorly understood. We have investigated the importance of methionine acquisition and synthesis for Streptococcus pneumoniae growth and virulence using strains with gene deletions affecting a putative methionine ABC transporter lipoprotein (Sp_0149, metQ) and/or methionine biosynthesis enzymes (Sp_0585 - Sp_0586, metE and metF). Immunoblot analysis confirmed MetQ was a lipoprotein and present in all S. pneumoniae strains investigated. However, vaccination with MetQ did not prevent fatal S. pneumoniae infection in mice despite stimulating a strong specific IgG response. Tryptophan fluorescence spectroscopy and isothermal titration calorimetry demonstrated that MetQ has both a high affinity and specificity for L-methionine with a K(D) of ∼25 nM, and a ΔmetQ strain had reduced uptake of C(14)-methionine. Growth of the ΔmetQ/ΔmetEF strain was greatly impaired in chemically defined medium containing low concentrations of methionine and in blood but was partially restored by addition of high concentrations of exogenous methionine. Mixed infection models showed no attenuation of the ΔmetQ, ΔmetEF and ΔmetQ/ΔmetEF strains in their ability to colonise the mouse nasopharnyx. In a mouse model of systemic infection although significant infection was established in all mice, there were reduced spleen bacterial CFU after infection with the ΔmetQ/ΔmetEF strain compared to the wild-type strain. These data demonstrate that Sp_0149 encodes a high affinity methionine ABC transporter lipoprotein and that Sp_0585 - Sp_0586 are likely to be required for methionine synthesis. Although Sp_0149 and Sp_0585-Sp_0586 make a contribution towards full virulence, neither was essential for S. pneumoniae survival during infection.
Conflict of interest statement
Figures
References
-
- Lau GW, Haataja S, Lonetto M, Kensit SE, Marra A, et al. (2001) A functional genomic analysis of type 3 Streptococcus pneumoniae virulence. Mol Microbiol 40: 555–571. - PubMed
-
- Darwin AJ (2005) Genome-wide screens to identify genes of human pathogenic Yersinia species that are expressed during host infection. Curr Issues Mol Biol 7: 135–149. - PubMed
-
- Mei JM, Nourbakhsh F, Ford CW, Holden DW (1997) Identification of Staphylococcus aureus virulence genes in a murine model of bacteraemia using signature-tagged mutagenesis. Mol Microbiol 26: 399–407. - PubMed
-
- Himpsl SD, Lockatell CV, Hebel JR, Johnson DE, Mobley HL (2008) Identification of virulence determinants in uropathogenic Proteus mirabilis using signature-tagged mutagenesis. J Med Microbiol 57: 1068–1078. - PubMed
-
- Brown JS, Gilliland SM, Holden DW (2001) A Streptococcus pneumoniae pathogenicity island encoding an ABC transporter involved in iron uptake and virulence. Mol Microbiol 40: 572–585. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
