

Requirement for Dlgh-1 in planar cell polarity and skeletogenesis during vertebrate development
- PMID: 23349879
- PMCID: PMC3551758
- DOI: 10.1371/journal.pone.0054410
Requirement for Dlgh-1 in planar cell polarity and skeletogenesis during vertebrate development
Abstract
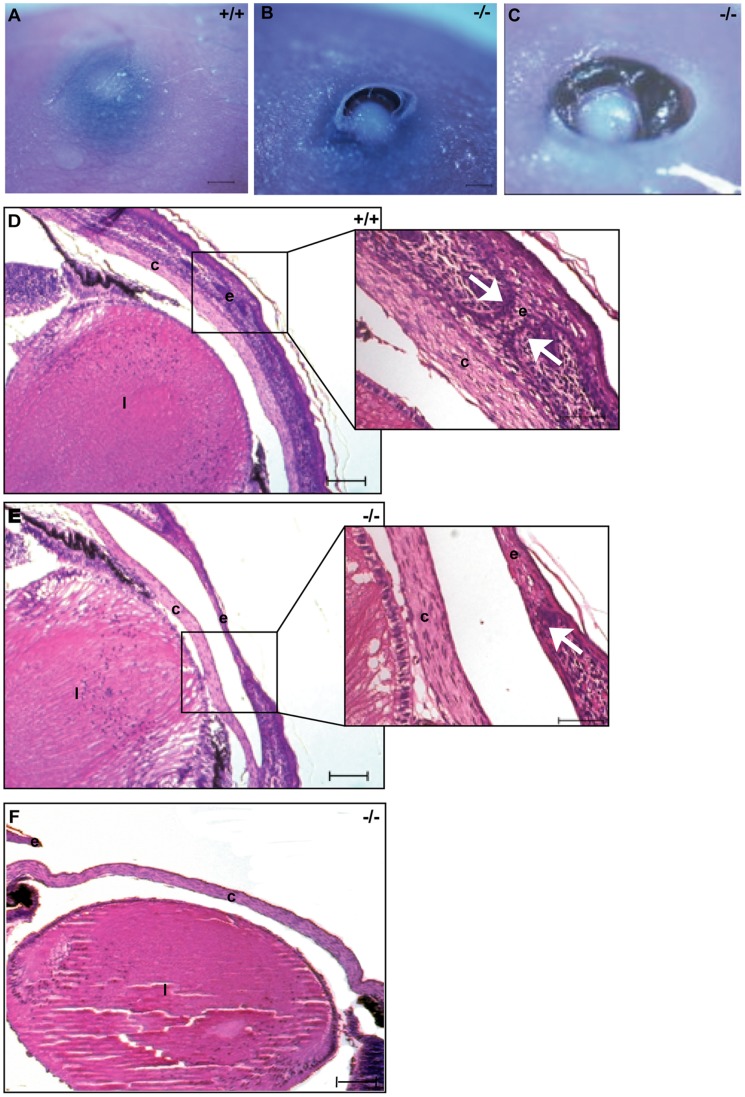
The development of specialized organs is tightly linked to the regulation of cell growth, orientation, migration and adhesion during embryogenesis. In addition, the directed movements of cells and their orientation within the plane of a tissue, termed planar cell polarity (PCP), appear to be crucial for the proper formation of the body plan. In Drosophila embryogenesis, Discs large (dlg) plays a critical role in apical-basal cell polarity, cell adhesion and cell proliferation. Craniofacial defects in mice carrying an insertional mutation in Dlgh-1 suggest that Dlgh-1 is required for vertebrate development. To determine what roles Dlgh-1 plays in vertebrate development, we generated mice carrying a null mutation in Dlgh-1. We found that deletion of Dlgh-1 caused open eyelids, open neural tube, and misorientation of cochlear hair cell stereociliary bundles, indicative of defects in planar cell polarity (PCP). Deletion of Dlgh-1 also caused skeletal defects throughout the embryo. These findings identify novel roles for Dlgh-1 in vertebrates that differ from its well-characterized roles in invertebrates and suggest that the Dlgh-1 null mouse may be a useful animal model to study certain human congenital birth defects.
Conflict of interest statement
Figures






References
-
- Bilder D, Li M, Perrimon N (2000) Cooperative regulation of cell polarity and growth by Drosophila tumor suppressors. Science 289: 113–116. - PubMed
-
- Wang Y, Nathans J (2007) Tissue/planar cell polarity in vertebrates: new insights and new questions. Development 134: 647–658. - PubMed
-
- Montcouquiol M, Rachel RA, Lanford PJ, Copeland NG, Jenkins NA, et al. (2003) Identification of Vangl2 and Scrb1 as planar polarity genes in mammals. Nature 423: 173–177. - PubMed
-
- Bossinger O, Klebes A, Segbert C, Theres C, Knust E (2001) Zonula adherens formation in Caenorhabditis elegans requires dlg-1, the homologue of the Drosophila gene discs large. Dev Biol 230: 29–42. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
