

Design principles of regulatory networks: searching for the molecular algorithms of the cell
- PMID: 23352241
- PMCID: PMC3664230
- DOI: 10.1016/j.molcel.2012.12.020
Design principles of regulatory networks: searching for the molecular algorithms of the cell
Abstract
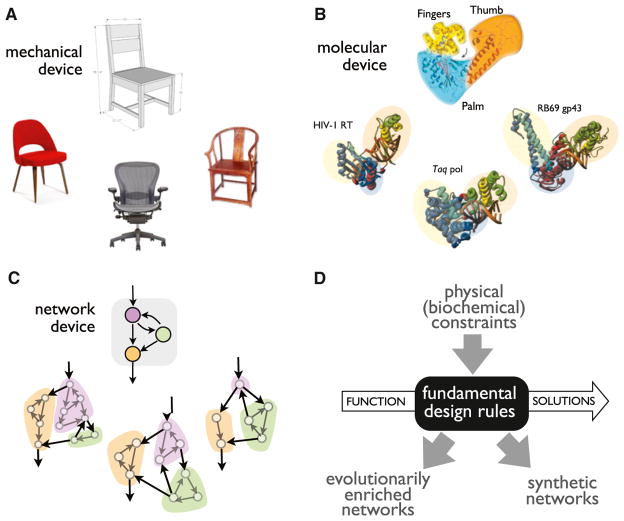
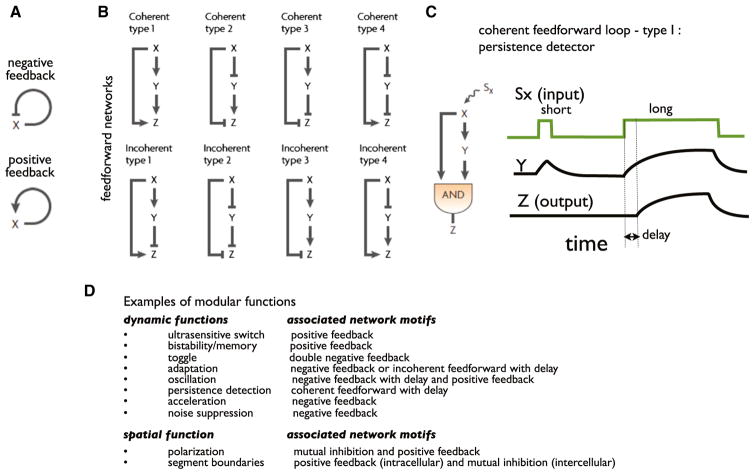
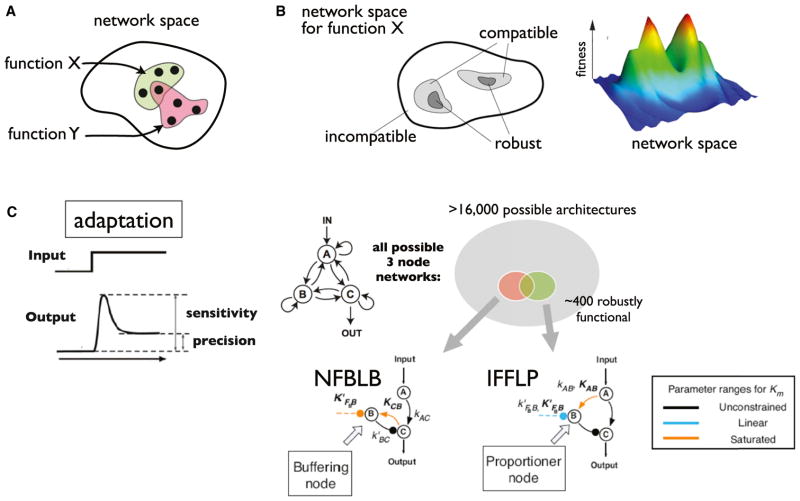
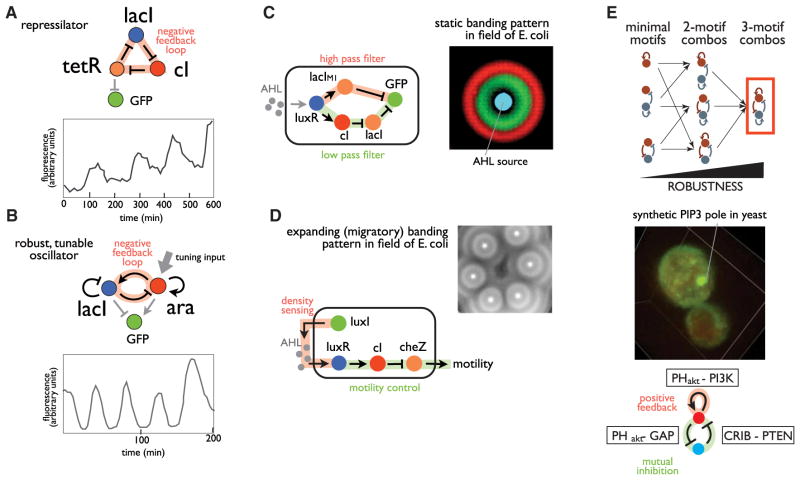
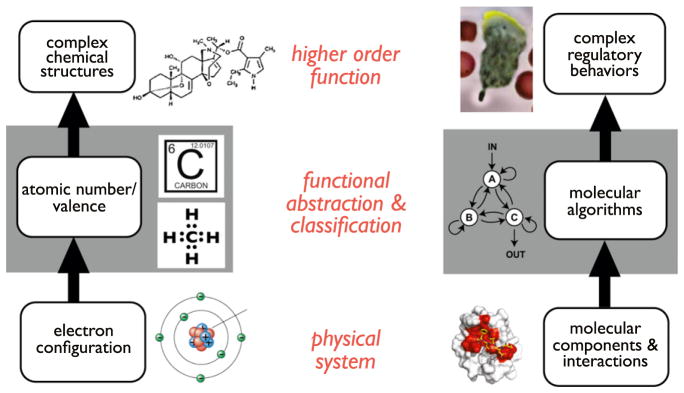
A challenge in biology is to understand how complex molecular networks in the cell execute sophisticated regulatory functions. Here we explore the idea that there are common and general principles that link network structures to biological functions, principles that constrain the design solutions that evolution can converge upon for accomplishing a given cellular task. We describe approaches for classifying networks based on abstract architectures and functions, rather than on the specific molecular components of the networks. For any common regulatory task, can we define the space of all possible molecular solutions? Such inverse approaches might ultimately allow the assembly of a design table of core molecular algorithms that could serve as a guide for building synthetic networks and modulating disease networks.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
A paradigm shift in medicine: A comprehensive review of network-based approaches.Biochim Biophys Acta Gene Regul Mech. 2020 Jun;1863(6):194416. doi: 10.1016/j.bbagrm.2019.194416. Epub 2019 Aug 2. Biochim Biophys Acta Gene Regul Mech. 2020. PMID: 31382052 Review.
-
Identification of functional modules using network topology and high-throughput data.BMC Syst Biol. 2007 Jan 26;1:8. doi: 10.1186/1752-0509-1-8. BMC Syst Biol. 2007. PMID: 17408515 Free PMC article.
-
Network legos: building blocks of cellular wiring diagrams.J Comput Biol. 2008 Sep;15(7):829-44. doi: 10.1089/cmb.2007.0139. J Comput Biol. 2008. PMID: 18707557
-
Synthetic biological networks.Rep Prog Phys. 2013 Sep;76(9):096602. doi: 10.1088/0034-4885/76/9/096602. Epub 2013 Sep 4. Rep Prog Phys. 2013. PMID: 24006369 Review.
-
Biological impacts and context of network theory.J Exp Biol. 2007 May;210(Pt 9):1548-58. doi: 10.1242/jeb.003731. J Exp Biol. 2007. PMID: 17449819 Review.
Cited by
-
Network representations and methods for the analysis of chemical and biochemical pathways.Mol Biosyst. 2013 Sep;9(9):2189-200. doi: 10.1039/c3mb70052f. Mol Biosyst. 2013. PMID: 23857078 Free PMC article. Review.
-
The Topological Characteristics of Biological Ratio-Sensing Networks.Life (Basel). 2023 Jan 28;13(2):351. doi: 10.3390/life13020351. Life (Basel). 2023. PMID: 36836707 Free PMC article.
-
Controlling Stochasticity in Epithelial-Mesenchymal Transition Through Multiple Intermediate Cellular States.Discrete Continuous Dyn Syst Ser B. 2016 Sep;21(7):2275-2291. doi: 10.3934/dcdsb.2016047. Discrete Continuous Dyn Syst Ser B. 2016. PMID: 29497351 Free PMC article.
-
Intracellular Information Processing through Encoding and Decoding of Dynamic Signaling Features.PLoS Comput Biol. 2015 Oct 22;11(10):e1004563. doi: 10.1371/journal.pcbi.1004563. eCollection 2015 Oct. PLoS Comput Biol. 2015. PMID: 26491963 Free PMC article.
-
MiRNAs confer phenotypic robustness to gene networks by suppressing biological noise.Nat Commun. 2013;4:2364. doi: 10.1038/ncomms3364. Nat Commun. 2013. PMID: 24077216 Free PMC article.
References
-
- Alon U. Network motifs: theory and experimental approaches. Nat Rev Genet. 2007;8:450–461. - PubMed
-
- Atkinson MR, Savageau MA, Myers JT, Ninfa AJ. Development of genetic circuitry exhibiting toggle switch or oscillatory behavior in Escherichia coli. Cell. 2003;113:597–607. - PubMed
-
- Aubel D, Fussenegger M. Watch the clock-engineering biological systems to be on time. Curr Opin Genet Dev. 2010;20:634–643. - PubMed
-
- Bar-Even A, Flamholz A, Noor E, Milo R. Rethinking glycolysis: on the biochemical logic of metabolic pathways. Nat Chem Biol. 2012;8:509–517. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
