

A common mode of recognition for methylated CpG
- PMID: 23352388
- PMCID: PMC3608759
- DOI: 10.1016/j.tibs.2012.12.005
A common mode of recognition for methylated CpG
Abstract
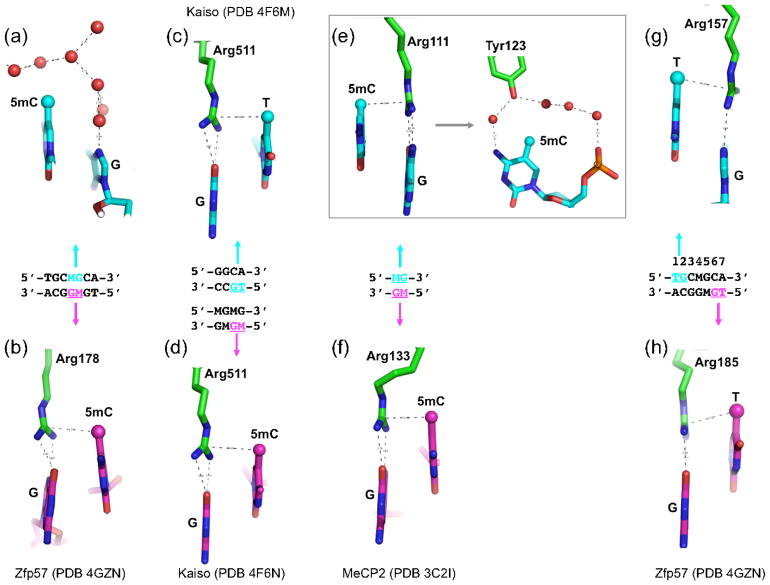
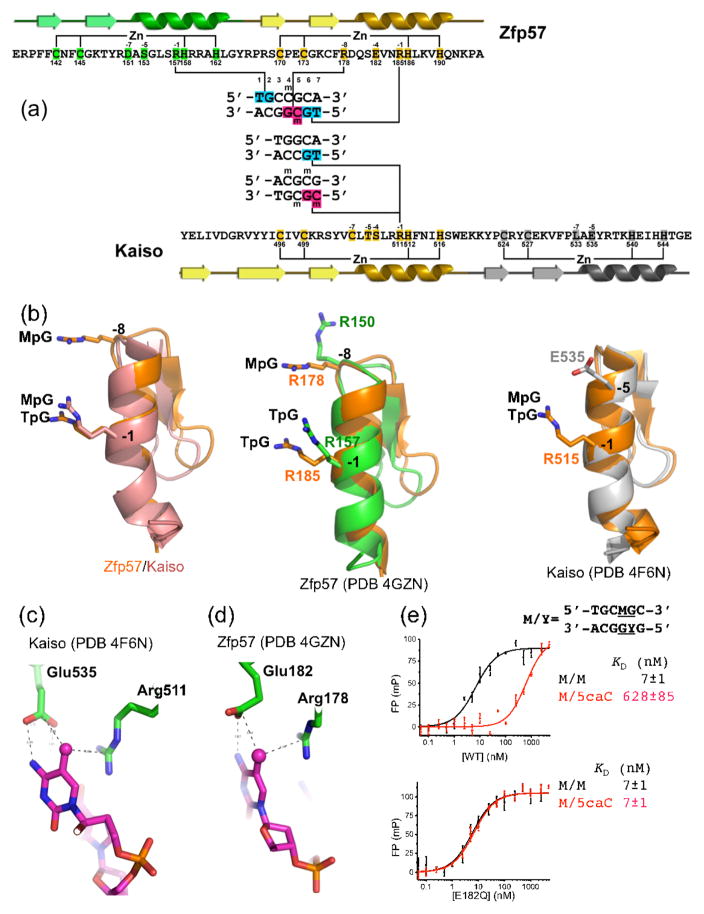
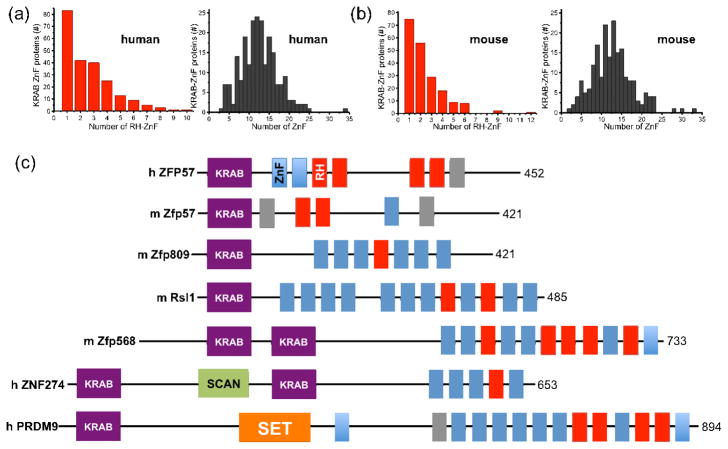
Much is known about vertebrate DNA methylation, however it is not known how methylated CpG within particular sequences is recognized. Two recent structures of C2H2 zinc finger (ZnF) proteins in complex with methylated DNA reveal a common recognition mode for 5-methylcytosine (5mC) that involves a 5mC-Arg-G triad. In the two ZnF proteins, an arginine that precedes the first Zn-binding histidine (RH motif) can interact with a 5mCpG or TpG dinucleotide. Among a family of >300 human Krüppel-associated box (KRAB) domain containing ZnF proteins examined, two-thirds contained at least one ZnF that included an RH motif. We propose that the RH-ZnF motifs provide specificity for 5mCpG, whereas the neighboring Zn fingers recognize the surrounding DNA sequence context.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Is there a telltale RH fingerprint in zinc fingers that recognizes methylated CpG dinucleotides?Trends Biochem Sci. 2013 Sep;38(9):421-2. doi: 10.1016/j.tibs.2013.06.005. Trends Biochem Sci. 2013. PMID: 23992945 No abstract available.
-
Response to mackay et Al.Trends Biochem Sci. 2013 Sep;38(9):423. doi: 10.1016/j.tibs.2013.06.010. Trends Biochem Sci. 2013. PMID: 23992946 No abstract available.
References
-
- Bestor T, et al. Cloning and sequencing of a cDNA encoding DNA methyltransferase of mouse cells. The carboxyl-terminal domain of the mammalian enzymes is related to bacterial restriction methyltransferases. J Mol Biol. 1988;203:971–983. - PubMed
-
- Okano M, et al. Cloning and characterization of a family of novel mammalian DNA (cytosine-5) methyltransferases. Nat Genet. 1998;19:219–220. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
